Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Май қышқылдарының алмасуыСодержание книги
Поиск на нашем сайте
Дәрістің мақсаты: Студенттерді липидтер алмасуы, олардың жалпы сипаттамасы, синтезі және реттелуі процестерінің механизмдерімен, олардың медициналық маңызымен таныстыру Дәріс жоспары:
1.Май қышқылдарының алмасуы. 2.Бета-тотығу бұл май қышқылдары катаболизмі үшін спецификалық жол. 3.Май қышқылдары катаболизмінің физиологиялық маңызы. 4.Май қышқылдарының биосинтезі. 5.Ацетосірке қышқылдардың түзілуі және қолданылуы, кетондық денелердің синтезі.Бұл процестің физиологиялық маңызы. 6.Стероидтар алмасуы. Холестерин басқа да стероидтар қатарының алғызаты ретіндегісі. 7.Холестериннің биосинтезі туралы түсінік. Холестериннің ағзадан шығарылуы. 8.Липидтер алмасуындағы бұзылулар.
Әдебиеттер: Негізгі: 1.Биохимия. Учебник /под редакцией член – корр РАН, проф. Е.С. Северина – М.: ГЭОТАР-МЕД, 2003.- 784 с 2. Биохимия. Краткий курс с упражнениями и задачами /под редакцией член – корр РАН, проф. Е.С. Северина и проф. А.Я. Николаева – М.: ГЭОТАР-МЕД, 2001.- 448 с 3. Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия человека: В 2-х томах. Т. 2. Пер. с англ: - М.: Мир, 1993. - 384с. 4.Николаев А.Я. Биологическая химия – М.: Высшая школа, 1989. - 494с.
Қосымша: 1.Бышевский А.Ш., Терсенов О. А. Биохимия для врача – Екатеринбург: Уральский рабочий, 1994. - 384с. 2. Гринстейн Б., Гринстейн А. Наглядная биохимия: Пер. с анг.-М.: ГЭОТАР-МЕД, 2000. - 119с 3. Тапбергенов С. О. Медицинская биохимия. – Астана, 2001. – 292с. 4. Эллиот В., Эллиот Д. Биохимия и молекулярная биология. – М. Мир – 1999.- 327с.
Майлардың катаболизмінде келесі процестер өтеді: 1.Липолиз (негізінен май ұлпасында); 2.Май қышқылдарының β-тотығуы (бауырда максимальды жылдамдықпен); 3.Ацетил-КоА кетондық денелерге айналып, басқа ұлпаларда жұмсалады; 4.Майлардың катаболизмі әртүрлі мүшелер мен ұлпаларда шоғырланған.
Май қышқылдарының алмасуы Май қышқылдары реакцияға тек активтенген соң ғана түсе алады. Активтенуі цитоплазмада өтіп, метаболизмі екі жолмен жүреді: 1.Ацетил-КоА-ға дейінгі катаболизмі (β-тотығу); 2.Майлар мен басқа липидтердің синтезі.
1. Май қышқылдарының алмасуы. Бета-тотығу май қышқылдарының катаболизмінің спецификалық жолы. Май қышқылдарының катаболизмінің физиологиялық маңызы. Қаннан жасушаға өткен май қышқылдары ацетил-КоА-синтетаза ферменті әсерімен активтелу реакциясына түседі.
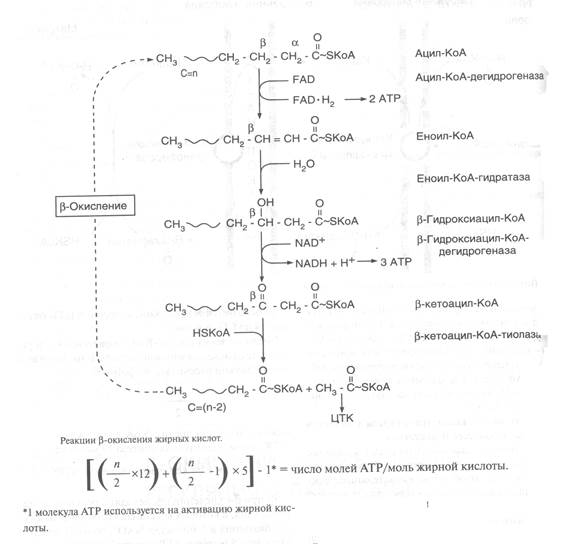
β-тотығу – бұл май қышқылдарының ацетил-КоА ыдырауының спецификалық жолы. Тотығу реакциясы β-көміртегі атомында жүретін болғандықтан β-тотығу деп аталады. Β-тотығу және одан түзілген ацетил-КоА 4КҚЦ-да тотығып, АТФ синтезі қызметін атқарады. β-тотығу процесі митохондрия матриксінде тек аэробтық жағдайда өтеді, себебі, ҮКҚЦ-мен байланысты. Митохондрияның ішкі мембранасынан ацетил-КоА өте алмайды, сондықтан май қышқылдарын тасымалдайтын корнитинация трансферазалық комплекске жұмыс істейді. Митохондрияның сыртқы мембранасында корнитинацил-трансфераза І ферменті ацилдік қалдықтың корнитинге ауысуын катализдейді. Кіші молекулалы ацилкарнитин транслоказа арқылы митохондрияның ішкі мембранасына өтеді. Оның бетіндегі корнитинациятрансфераза ІІ ацилдік қалдықты митохондрияішілік HS-КоА-ға ауысуын катализдейді. Нәтижесінде, ацетил-КоА түзіледі.
Сыртқы мембрана Ішкі мембрана Цитозол
Т Р А Н С Л О К А З А
 матрикс матрикс
// О
Сур. 9. Май қышқылдарының мембрана арқылы тасымалдануы.
Май қышқылының тотығуында түзілетін АТФ мөлшерін келесі теңдеумен есептеуге болады:
АТФ саны = [{ (n/2) x 12} + { (n/2 –1) } x 5] – 1*
Бұл жердегі n – көміртегі саны; 1* - ацил-КоА-да жұмсалатын АТФ. Мысалы, пальмитин қышқылы (С16) тотыққанда, β-тотығудың 7 циклі өтеді. Нәтижесінде, 8 моль ацетил-КоА, 7 моль ФАДН2, 7 моль НАДН+Н+ түзіледі. Сондықтан β-тотығуда 35 АТФ, ал 4 КҚЦ-да 96 АТФ, барлығы 131 АТФ молекуласы түзіледі.
b-тотығудың реттелуі.b-тотығудың жылдамдығы, басқа да метаболиттік жолдар тәрізді, ацил-КоА субстратының жеткілікті болуына тәуелді болады. Сондықтан, май қышқылдарының b-тотығуы постабсорбтық кезеңде немесе ұзақ мерзімді физикалық жұмыс кезінде май ұлпасындағы майлардың ыдрауы нәтижесінде қанда май қышқылдарының концентрациясының жоғарлауында ол активтенеді. Бұл кезеңдерде бұлшықеттер, миокард және бауыр май қышқылдарын энергия көзі ретінде белсенді түрде пайдаланады. Ми май қышқылдарын энергия көзі ретінде пайдаланбайды, себебі гидрофобты молекула болғандықтан гематоэнцефольдық бөгет арқылы өте алмайды.
Май қышқылдарының b-тотығуы b-тотығудың реттеуші ферменті - карнитинацилтрансфераза I. Бұл ферменттің аллостерлік ингибиторы – малонил-КоА. Ингибитор тек май қышқылдарының биосинтезінде ғана түзіледі. Сондықтан, постабсорбтық кезеңде, бұл ацетильдік қалдықтардың митохондриядан цитозолға түсуінің тоқтатылған кезі, малонил-КоА-ның да синтезі тежеледі және β-тотығу бұл ингибитор болмағандықтан белсенді жүреді. АТФ-ті беретін маңызды жол болғандықтан β-тотығу жасушада энергия қажеттілігі жоғарлаған кезде активтеледі. Бұл β-тотығу реакцияларының электрон тасымалдаушы тізбекпен (ЭТТ) NAD және FAD коферменттері арқылы тікелей байланысатындығынан мүмкін болады.АТФ-тің жұмсалуы интенсивті болған сайын май қышқылдарының тотығуы да жылдамдайды. Себебі, жаңа АТФ молекулаларының синтезін қамтамассыз ету қажет болады. 2. Май қышқылдарының биосинтезі. Май қышқылдарының синтезі абсорбтық кезеңде, қандағы глюкоза концентрациясы жоғарлаған кезде, бауырда және май ұлпаларының цитоплазмасындағы эндоплазмалық ретикулумның мембранасында жүреді. Бұл кезеңде гликолиз бен глюкозаның пентозофосфаттық ыдырау жолдары активтенеді. Соның нәтижесінде май қышқылдарының синтезі үшін қажетті субстраттар - ацетил -КоА, NADPH2, АТФ-тар түзіледі. Ацетил-КоА-ның пируваттың тотығып декарбоксилденуі нәтижесінде түзілуі митохондрия матриксінде өтеді, бірақ ацетил-КоА үлкен молекула болғандықтан митохондрия мембранасы арқылы май қышқылдарының синтезі өтетін цитоплазмаға түсе алмайды. Сондықтан ацетил-КоА оксалоацетатпен конденсацияланып цитратқа айналады және бұл түзілген цитрат транслоказа көмегімен ғана цитоплазмаға тасымалданады.
Цитоплазмада цитратлиаза ферменті әсерінен келесі реакция жүреді: Цитрат + HSKoA + АТР ® Ацетил-КоА + ADP + Рi + Оксалоацетат Цитоплазмаға ауыстырылған Ацетил-КоА май қышқылдарының синтезі үшін бастапқы субстрат болып табылады. Оксалоацетат цитозолда келесі түрленуге түседі: Оксалоацетат ® малат ® пируват. Соңғы реакция малик-ферментпен катализделінеді. Пируват қайтадан митохондрия матриксіне ауысады, ал малик-фермент әсерінен тотықсызданған NADPH2 сутегі доноры ретінде келесі май қышқылының синтезі үшін пайдаланылады. NADPH2 —нің басқа шығу көзіне пентозофосфаттық жол жатқызылады. Май қышқылдары синтезінің бірінші реакциясы — ацетил-КоА-ны малонил-КоА-ға айналдыру. Бұл реакцияны катализдейтін фермент,ацетил-КоА-карбоксилаза, май қышқылдары биосинтезіндегі реттеуші фермент болып табылады. Ол лигазалар класына жатқызылады, коферменті – биотин. Май қышқылдары синтезінің келесі реакциялары майқышқылдары синтазасынемесе пальмитатсинтаза деп аталатын ферментарқылы атқарылады. Себебі адам липидтерінің негізгі май қышқылы пальмитин қышқылы болып табылады. Ағзадағы май қышқылдарының синтезі пальмитин қышқылынан бастап түзіледі. Майқышқылы синтазасы эндоплазмалық ретикулум мембранасына бекітілген фермент. Ол полиқызметті фермент, екі идентті полипептидтік тізбектен тұрады. Олардың әрқайсысында 7 активті орталық және ацилтасымалдаушы белоктары болады. Ацилтасымалдаушы полипептидтік тізбек синтез барысында ұзарушы май қышқылы тізбегін бір активті орталықтан келесіне тасымалдайды. Әрбір белокта құрамында SH-тобы болатын екі байланыстырушы орталықтар болады. Тек малонил-КоА-ны байланыстыратын аймақтың құрамында белок емес компонент – B3 витамині (пантотен қышқылы) болады. Май қышқылдарының синтезі бастапқыда ацетилдік сонан кейін малонилдік қалдықты ацетитрансфераза және малонилтрансфераза көмектерімен майқышқылдары синтазасына ауыстырумен басталады. Ары қарай малонилдің карбоксилдік тобы СО2 түрінде бөлінеді және босатылған валентті орынға ацетил байланысып ацетоацетил-Е түзіледі. Келесі өтетін тотықсыздану, дегидраттану және тағыда тотықсыздану реакцияларының нәтижесінде бутирилдің радикалының түзілуі жүреді. Ол ферментпен байланысқан қалыпта болады. Сонан кейін тағыда осындай реакциялар циклі өтеді де соның нәтижесінде 6 көміртегі атомынан тұратын май қышқылының радикалы түзіледі. Бұл циклдер ары қарай пальмитин қышқылы түзілгенге дейін қайталанады. Осыдан кейін ғана, қажет болған жағдайда тиоэстераза ферменті май қышқылын майқышқылдары ферментінен бөліп тастайды.
Май қышқылдарының биоситезі бірегей реакциялардың қайталануы болып табылады. Сондықтан, бұл процесс циклдік деп аталады және әрбір цикл сайын май қышқылының радикалы екі көміртегі атомына ұзарады. Оның шығу көзі малонил-КоА болып табылады. Әрбір циклде тотықсыздану реакциясы NADPH+Н+ -ты пайдалану арқылы өтеді, ал оның шығу көзі глюкозаның пентозофосфаттық тотығу және және малик-ферментпен өтетін каталиттік жолдары болып табылады. Адам ағзасындағы басқа май қышқылдары пальмитин қышқылынан синтезделеді. Бұл кезде көміртектік тұлғаның малонил-КоА-ны және дегидрлену реакциясын пайдаланып ұзаруы жүреді. Адам ағзасында С9 көміртегі атомынан ары қарай қос байланысы қалыптасатын қанықпаған май қышқылдары синтезделмейді. Сондықтан адам ауыстырылмайтын май қышқылдарын тағам құрамынан алуы қажет. Қанықпаған май қышқылдары десатураза және басқа да ферменттер қатысуымен күрделі реакциялар арқылы микросомада синтезделеді. Қаныққан май қышқылдарының синтезі негізінен митохондрияда өтеді. Синтезделген май қышқылдары бос күйінде қалмайды, керісінше тез арада майлардың синтезіне және аз мөлшерде фосфолипидтердің синтезіне жұмсалады. Май қышқылдарының синтезінің жылдамдығы реттеуші фермент ацетил-КоА-карбоксилазаның активтілігіне тәуелді. Бұл ферменттің активтілігі келесі жолдармен реттеледі: 1. аллостерлік жолмен, цитрат активатор, ал пальмитоил–КоА – ингибитор болып табылады. 2. фосфорландыру – дефосфорландыру жолымен. Тағам қабылдаған соң инсулиннің әсерімен фосфатаза активтелінеді, ал ол ацетил-КоА-карбоксилазаны дефосфорланған активті формаға әкеледі. Бұл форма аллостерлік реттелуге түседі. Аштық және физикалық жұмыс кезінде глюкагон немесе адреналин аденилатциклаздық жүйе арқылы ацетил-КоА–карбоксилазаны фосфорланған активсіз формаға әкеледі. (Сур.14). Инсулин май қышқылдарының синтезіне қатысатын глюкозаның ыдырау өнімдері түзілуін катализдейтін басқа ферменттердің синтездеріне де ықпал етеді. Жоғары май қышқылдарының синтезі жүруі үшін қажетті жағдайлар:
1) Көмірсулардың түсуі, олар тотыққан кезде қажетті субстраттар мен NADPH+Н+ түзіледі. 2) Жасушаның жоғары энергиялық қабілеті – АТФ құрамының жоғары болуы. Ол цитраттың митохондриядан цитоплазмаға өтуін қамтамассыз етеді. Көмірсулар алмасуы және майлар алмасуы өзара тығыз байланысты болады. Көмірсулар майларға оңай айналады, ал майлардың көмірсуларға тікелей айналуы мүмкін емес. Себебі май қышқылдарынан түзілетін ацетил-КоА пируватқа айнала алмайды. Майлар мен көмірсулардың алмасулары энергия алмасуы ретінде біріктіріледі және ол гормондардың бақылауында болады. 3. Ацетосірке қышқылының түзілуі және қолданылуы, кетондық денелердің синтезі. Бұл процестің физиологиялық маңызы. Бауырда май қышқылдарының бір бөлігі кетондық денелерге түрленеді. Қалыпты жағдайда олардың қандағы концентрациясы аса жоғары емес - 1-3 мг/дл. Кейбір жағдайларда олардың синтезі жоғарлайды. Кетондық денелер – ацетоацетат және b-гидроксибутират – кейбір жағдайларда энергия көзінің қызметін атқарады. Бұл жағдайлар глюкозаны пайдаланудың мүмкіндігі төмендеген кезде (мысалы, аштықта, постабсорбтық кезеңде, құрамында көмірсуы аз, ал майлары көп тағамды қабылдау кезінде, ұзақ мерзімді физикалық жұмыс кезінде, қант диабетінде) туындайды. Кетондық денелер гематоэнцефалдық бөгет арқылы өте алады, себебі, гидрофилді молекула болып табылады. Бұлшықет пен бүйректер кетондық денелерді оның қандағы концентрациясы төмен жағдайында да энергия көзі ретінде пайдаланады. Кетондық денелердің синтезі бауыр жасушаларының митохондрияларында жүреді. Синтездің бастапқы субстраты ацетил-КоА b-тотығу процессі нәтижесінде түзіледі. (Сур.11). Аштық кезінде глюкагон гормоны аденилатциклаздық жүйе арқылы май ұлпасында майлардың ыдырауын активтендіреді. Май қышқылдары қанға бөлінеді және альбуминмен комплекс түзіп бауырға тасымалданады. Бауырда β-тотығудың жылдамдығы күшейеді, соның нәтижесінде ацетил-КоА-ның көп мөлшері түзіледі. Бірақ ацетил-КоА-ның Кребс цикліндегі тотығу жылдамдығы төмен болады, себебі, цитрат циклінің реттеуші ферментінің активтілігі төмен, өйткені оның аллостерлік ингибиторлары АТФ пен NADH-тың концентрациясы β-тотығу нәтижесінде жоғарлайды. Мұнан басқа, NADH-тың жоғары концентрациясында оксалоацетат малатқа дейін тотықсызданады және осы формада цитозолға тасымалданады, ол жерде реакция кері бағытта жүреді, сондықтан оксалоацетат глюконеогенездің субстратына айналады. Нәтижесінде, митохондрияда ацетил-КоА жиналып қалады және ол кетондық денелердің синтезіне жұмсалады. Адамда негізгі кетондық денеге β-гидроксибутиратты жатқызады. Себебі митохондриядағы NADH-тың жоғары концентрациясы жоғарлауы жағдайы ацетоацетаттың тез тотықсыздануын қамтамассыз етеді. Ұзақ ашыққанда және қант диабетінде ацетоацетаттың қандағы концентрациясы біршама айқын жоғарлайды. Бұл жағдайда ацетоацетат ферментсіз түрде декарбоксилденіп үшінші кетондық дене ацетонға айналады. Ацетон ағзада энергия көзі ретінде жұмсалмайды. Сондықтан зәр, тер және шығарылған дем құрамдарында ағзадан шығарылады. Бұрынғы кезде гипергликемиялық комадағы науқасқа ауызынан шыққан ацетонның исі бойынша («шіріген алма иісі») қант диабетінің диагнозын қойған болатын. 4.Стероидтар алмасуы. Холестериннің кейбір стероидтардың алғызаты екендігі. Холестериннің биосинтезі туралы түсінік. Холестериннің ағзадан шығарылуы жолдары. Холестерин ағзадағы басқа да кейбір стероидтардың - өт қышқылдарының, стероидтық гормондардың және D3 – витамині синтездерінің алғызаты болып табылады. Холестерин барлық жасушалардың мембранасы құрамының құрылымдық компоненті болып табылады. Холестерин ағзаға екі жолмен түседі: 1. жануар тектес тағамдар арқылы (экзогендік холестерин) және бауырда синтезделуі арқылы (эндогендік холестерин). Холестерин аз мөлшерде бауырдан басқа мүшелерде - ішек пен тері жасушаларында да синтезделе алады. Ағзада холестерин екі қызметті атқарушы әртүрлі фондыны құрайды. Олардың арасында тұрақты түрдегі алмасу болады. Бос холестерин фонды (бірінші фонд) – бұл ең үлкен фракция болып табылады. Оның құрамына мембрана холестерині, қандағы липопротеиндердің моноқабатты фосфолипидтік холестерин кіреді. Холестерин эфирлерінің фондысы (екінші фонд) – бұл толық гидрофобты ағзаның резервтік холестериндік формасы болып табылады. Холестериннің бұл формасы жасуша цитозолының липидтік тамшылары және липопротеиндердің ядролары құрамдарында анықталған. Адам ағзасындағы холестериннің жалпы мөлшері 140 г, 93%-ы ұлпаларда кездеседі. Көптеген мүшелерде холестериндік құрам 100 г ұлпаға 0,1 – 0,3 г –нан келеді. Тек жүйке жүйесінің ұлпасы 100 г-на 2 г және бүйрек үстілер 100 г-на 10 г холестериннен келуі бойынша ерекшелінеді. Холестериннің 80%-ға жуығы жүйке, бұлшықет, дәнекер және май ұлпаларында шоғырланған. Жасушада холестерин келесі түрде таралған: - плазмалық мембранада холестерин/фосфолипидтің молярлық қатынастары орташа есеппен 1:1 құрайды немесе фосфолипипдтің бір молекуласына холестериннің бір молекуласы келеді, сонымен бірге биқабатты мембрананың сыртқы қабатында 2/3, ал ішкі қабатында 1/3 холестериндік қатынас қалыптасқан. Жасушаішілік мембранадағы холестерин құрамы плазмалық мембранамен салыстырғанда 10 есе аз болады. Жалпы холестериннің 7—10%-ы липопротеиндер құрамында қанда және лимфада болады. Оның негізгі бөлігі (70%) холестерин эфирлері болып табылады. Жас ұлғайғанда мүшелердегі холестериннің мөлшері де ұлғая бастайды. Мысалы, сәбилерде (1 жас) қан плазмасындағы холестериннің мөлшері 50±10 мг/дл болса, ересек адамдарда 200±40 мг/дл, немесе 5,2±1,3 ммоль/л болады. Холестерин ағзадағы стероидтар – кортикостероидтардың, андрогендердің, эстрогендердің, өт қышқылдарының, D-витамині синтездерінің алғызаты болып табылады. бұл заттардың синтезі үшін тәулік сайын 0.5 – 0.7 г холестерин жұмсалады. Фекалий құрамында тәулігіне 1,3 г холестерин өт қышқылдары және өт қышқылы мицеллалары құрамындағы холестерин түрінде шығарылады. Тағамнан тек жануар өнімдері тектес түрлерінен ғана түскен холестерин қабылданады. Адамның асқазан – ішек трактісінде өсімдік тектес өнімдер құрамында кең таралған b-ситостерол - стероид практика жүзінде сіңірілмейді. Ағзаға тағам құрамында түскен холестериннің эфирлері панкреаттық немесе ішек сөлінің холестеразасы арқылы гидролизденеді. Гидролиз өнімдері қоспалы мицелла түрінде ішек эпителийлері арқылы сіңіріледі. Экзогендік немесе ішек жасушалары арқылы синтезделген холестерин жартылай түрде оның эфиріне айналады. Бұл процесс екі стадиядан тұрады: 1. май қышқылының ацил-КоА-синтетаза әсерінен активтендірілуі; 2. ацилдік қалдықты ацил-КоА-дан холестериннің НО-тобына қосу. Ол ацилхолестеролацилтрансфераза (АХАТ) ферменті арқылы катализденеді.
Сур. 2. Кетондық денелердің синтезі Ішектің шырышты қабатының жасушаларында холестерин эфирлері, бос холестерин, ресинтезделген ТАГ, және энтероциттерде синтезделген аполипопротеиндер В-48, A-I және A-II хиломикрондар құрамына жинақталады. Оллар алғашқыда лимфаға, сонан кейін қанға түседі. Қан ағынында хиломикрондар ТЖ ЛП-лармен жанасады және олардан белоктарды (апоС-П және апоЕ) алады. Кері бағытта хиломикрондардан ТЖ ЛП-ларға апоА-1 және апоА-II белоктары түседі. Бұл алмасудың нәтижесінде хиломикрондар липопротеинлипазамен байланысуға қабілетті жетілген бөлшектерге айналады. Хиломикрондар құрамына кіретін ТАГ қылтамыр (капилляр) қабырғасына бекітілген липопротеинлипаза арқылы бөлініп алынады, ал хиломикрондар қалдық хиломикрондарға айналады. Қалдық хиломикрондар қан ағынынан бауырда эндоцитоз механизмімен алынады, сонан кейі лизосомалық ферменттер арқылы ыдыратылады. Қалдық хиломикрондардан және басқа ЛП-лардан босатылған холестерин ағзадағы холестериннің жалпы фондысы құрамына кіреді. Ол сонымен бірге, бауырдағы эндогендік холестериннің және ТТ ЛП- рецепторлардың синтездерін төмендетеді. Ағза үшін маңызды болып табылатын холестерин синтездеуші ұлпалары – бауыр, ішектер, бүйрек үстілердің қыртысты қабаты, тері, репродуктивті мүшелер, плацента болып табылады. Бұл стероидтың 80%-ға жуығы бауырда түзіледі. Бауыр холестеринді тек өзінің қажеті үшін ғана пайдаланбайды, сонымен бірге басқа мүшелер мен ұлпаларға липопротеиндер құрамында тасымалдайды. Холестериннің биосинтезі, негізінен, бауырда, гепатоциттердіңэндоплавзмалық ретикулумінде өтеді (Сур.4). Бұл эндогендік холестерин болып табылады. Холестериннің бауырдан ұлпалараға тасымалдануы тұрақты түрде жүреді. Мембраналарды құрастыру үшін тағамдық немесе экзогендік холестерин де пайдаланылады. Холестериннің биосинтезінің негізгі ферменті - ГМГ-редуктаза (b-гидрокси, b-метил, глутарил-КоА редуктаза). Холестериннің биосинтезі процессін шартты түрде 3 этапқа бөлуге болады. (Сур.15). Мевалонаттың 3 ацетил-КоА қалдығынан түзілуі (І этап). Ацетил-КоА цитоплаззмаға цитрат түрінде жеткізіледі. Синтездің бастапқы реакцияларының кезектесулерінің реті кетондық денелер синтезі реакцияларына ұқсас, бірақ кетондық денелердің синтезі митохондрияда өтеді. Цитозолда алғашқы түзілген ГМГ-КоА-ның әрбір молекуласы ГМГ-КоА-редуктаза (b-гидрокси, b-метил, глутарил-КоА редуктаза) арқылы NADPH+H+-тың екі молекуласын пайдаланып мевалонатқа дейін тотықсызданады. Бұл реакция негізгі реттеуші болып табылады және берілген метаболиттік жолдың жылдамдығын шектейді.
Скваленнің мевалонаттың 6 молекуласынан түзілуі (ІІ этап). Бұл этаптың барысында мевалонат молекуласынан (мевалонаттың 1 молекуласына 3 АТФ молекуласы жұмсалып) фосфорланған 5-көміртекті изопроеноидтық туындылар – изопентенилпирофосфаттар түзіледі. Олардың конденсациясы 30 көміртек атомды қосылыс – скваленнің түзілуіне әкеледі.
|
|||||||||
|
|
Последнее изменение этой страницы: 2024-06-17; просмотров: 9; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.218.63.231 (0.016 с.) |

 RCOOH+HSKoA+ATP ® R-CO-SKoA+AMP+PPi
RCOOH+HSKoA+ATP ® R-CO-SKoA+AMP+PPi

 R-C-S-KoA корнитин R-C~KoA
R-C-S-KoA корнитин R-C~KoA
 || \\
|| \\ O корнитин- корнитин- О
O корнитин- корнитин- О ацилтрансфераза трансфераза ІІ
ацилтрансфераза трансфераза ІІ
 Малонил-КоА
Малонил-КоА


 HSKoA R-C-корнитин HSKoA
HSKoA R-C-корнитин HSKoA