
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Введение в количественную биологиюСодержание книги Поиск на нашем сайте
Введение в количественную биологию Васильев Алексей Артёмович при участии Окштейна Игоря Леонидовича (семинары в рамках лекционно-семинарского курса «Основы биологии») Часть 2 (весенний семестр) От молекулярного конструктора к процессам на уровне клетки и организма
Обозначения в ссылках: ТБл – Васильев А.А. «Теоретическая биология», часть 1, М., МФТИ, 2002. ФЖ – Шмидт-Ниельсен К. «Физиология животных: приспособление и среда», в 2-х т., М., Мир, 1982. РЖ – Шмидт-Ниельсен К. «Размеры животных: почему они так важны?», М., Мир, 1988. ОЖ – Шмидт-Ниельсен К. «Как работает организм животного», М., Мир, 1976. БфХ= «Биофизическая химия» – Кантор Ч., Шиммел П. «Биофизическая химия», т.3 ФГ = Федоров В.Д., Гильманов Т.Г. Экология. –– М.: Изд-во МГУ, 1980 «Биофизика» – Волькенштейн М.В. «Биофизика» М.: Наука, 1988
Введение. В ходе жизнедеятельности клетки совершаются многочисленные молекулярные процессы – биохимические превращения, перенос молекул разного размера и целых органоидов на разные расстояния, конформационные перестройки на уровне отдельных молекул и их комплексов, изменения свойств среды (закисление, изменение ионной силы растворов и осмотического давления, накопление органических веществ). Жизнедеятельность на молекулярном уровне (и соответствующую молекулярную активность) можно рассматривать как то, что в принципе может быть реализовано средствами молекулярного конструктора. отсюда, соответственно, расширение списка используемых в жизнедеятельности процессов и целесообразность существующих возможностей
В части 1 рассматривалось связывание фермента или активного макромолекулярного комплекса с несколькими одинаковыми молекулами (кооперативное связывание) как способ регуляции молекулярной активности. Получаемые свойства регулируемых процессов представляют кривые связывания – зависимости скорости процесса от концентрации молекул регулятора. Например, при кооперативном связывании регулятора (c) с ферментом (E) по схеме
при наличии 4-х центров связывания было получено выражение для относительной доли активной формы фермента, связанного с 4-мя молекулами регулятора (E c 4) Y = [E c 4]/ [E] + [E c ] + [E c 2] + [E c 3] + [E c 4] = = K4K3K2K1c4/(1 + K1c + K2K1c2+ K3K2K1c3+ K4K3K2K1c4), которая при малых концентрациях регулятора пропорциональна 4-й степени этого регулятора и выходит на насыщение при высоких концентрациях регулятора. В этом случае вторая производная Y’’ по концентрации регулятора изменяет знак с положительного на отрицательный. Говорят, что такая кривая имеет S-образный или сигмоидный вид (подобно наклоненной и вытянутой вправо латинской букве S).
Со свойствами кооперативных процессов молекулярного уровня непосредственно связано проявление жизнедеятельности на уровне клетки и организма в целом. Примеры связи молекулярных процессов и их характеристик (свойств) с явлениями свойствами на макроскопическом (физиологическом) уровне: – сложная функция связывания гемоглобина с О2, CО2 и другими молекулами (CО, ДФГ как регулятор) в процессах дыхания (БфХ, т.3; ФЖ, т.2) – актомиозин в мышечном сокращении (МБК, «Биофизика», ФЖ, т.2) – мембранные структуры и разнообразие процессов с их участием –и передача нервного импульса («Биофизика», термодинамические свойства мембраны в этой связи – Харакоз), прочность на уровне тканей (обсуждение процессов разрушения в этой связи – Партон «Механика разрушения) и т.д.
Из перечисленных процессов интересен транспорт кислорода в организме, который удобно сравнивать с регуляцией молекулярной активности, которая подробно обсуждалась в части 1. Для регуляции молекулярной активности, как правило, важно получить высокую чувствительность к некоторому сигналу или воздействию, а эффективность выражает полнота использования максимальных производительных возможностей при отсутствии паразитного фона в неактивном состоянии. Поэтому два состояния, между которыми нужно организовать переход (переключение) – это полное включение (одно состояние) и полное выключение (другое состояние). Иными словами, «идеальному переключателю» отвечает переход, близкий к ступенчатому, т.е. высокая чувствительность в заданном диапазоне, и кривую насыщения важно приблизить к кривой «идеального переключателя». При этом нежелателен и совсем резкий переход (ступенька, т.е. коэффициент усиления, стремящийся к бесконечности), т.к. тогда будет слишком высокая чувствительность к шуму.
Перенос кислорода кровью Целесообразность и необходимость значительного изменения насыщения в относительно узком диапазоне концентраций кислорода определена обычными условиями, в которых осуществляется дыхание животных. Для транспорта важен конечный эффект переноса на одну молекулу переносчика, который определяет разность принятого и отданного кислорода. А именно, количество перенесенного кровеносной системой и использованного кислорода дает баланс поступления кислорода в кровь и его отдачи в тканях, что можно представить/выразить как разность количеств кислорода на входе и на выходе из них
где W – объем, перекачиваемый кровеносной системой в единицу времени, c – концентрация гемоглобина в единице объема (с пересчетом в количество переносимого кислорода при насыщении гемоглобина), Y (P) – зависимость насыщения гемоглобина от парциального давления кислорода в равновесии с кровью при контакте с внешней средой в органах дыхания (на выходе артериальной крови из них, индекс «a») и в контакте с потребляющими тканями (на выходе из них, т.е. в венозной крови, индекс «v»). При этом от величин Pa и Pv зависят скорости поступления кислорода на других этапах (см. далее), их (величины Pa и Pv) естественно определить как верхнюю и нижнюю (Pa > Pv) рабочую точку на кривой насыщения гемоглобина соответственно.
В силу такой зависимости эффекта переноса (кислорода связанного с гемоглобином) от DY важно не только то, сколько кислорода содержит артериальная кровь, насыщаемая при контакте с кислородом из внешней среды в органах дыхания (ее определяет насыщение Y(Pa)), но и способность отдавать кислород при давлении в капиллярах. Если величина Y(Pv) велика, то кровь будет плохо отдавать кислород. Иными словами, для гемоглобина (как переносчика) желательно возможно б о льшее значение DY = Y(Pa) – Y(Pv) при обычных условиях.
При дыхании в воздухе значение Pa не больше парциального давления в воздухе (в действительности в легких парциальное давление кислорода составляет около 100 Торр, т.е. меньше парциального давления в воздухе 150 Торр). Значение Pv не может быть меньше критического значения Pv,cmin, которое должно обеспечить последующую диффузию из капилляров в клетки (см. ниже). При высокой интенсивности потребления последняя достаточно велика (см. расчет далее), а значит, большое значение DY желательно обеспечить при изменении давления от Pa до Pv, происходящем в относительно узком диапазоне. Эта ситуация очень похожа на требования к регуляции молекулярной активности с необходимостью переключения в заданном узком диапазоне. Причем в случае транспорта ограничения даже более жесткие, чем при регуляции молекулярной активности. При регуляции, по сути, происходит передача информации, т.е. интенсивность потока материального носителя сигнала (информации) может быть очень низкой. В случае потока низкой интенсивности затраты в абсолютном выражении малы. Регуляторную цепочку в принципе также можно дополнить еще одним этапом (или несколькими этапами), увеличивая усиление преобразуемого сигнала. Такое усложнение цепочки не будет обременительным с точки зрения затрат ресурсов или требований организационного характера. При необходимости промежуточные этапы усиления целесообразны, что делает возможным подбор ширины диапазона для отдельных этапов под возможности молекулярного конструктора. Напротив, при транспорте для потока высокой интенсивности возможности такого рода либо значительно увеличивают затраты на осуществление транспорта и общие затраты организма при встраивании более сложно организованного транспорта в систему физиологических процессов, либо исключены физически. Если нужно передать сигнал (т.е. информацию), то один носитель сигнала вполне можно заменить на другой, а если нужен кислород как окислитель, то его практически невозможно заменить (хотя молекулы кислорода можно к чему-то присоединить, объединить несколько молекул кислорода вместе и т.п.). Развитие: интересно попробовать обсудить возможности модификации последовательностей этапов для процессов регуляции и транспорта (предложить критерии оценки эффекта, затрат разного рода и т.д.) Таким образом, значения Pa и Pv, по сути, заданы (внешними условиями и другими особенностями физиологической организации жизнедеятельности) и нужно существенное изменение DY (желательно близкое к 1) в заданном узком диапазоне. Иными словами, «идеальный эффект» в данном случае аналогичен обсуждаемому выше при переключении молекулярной активности – это достаточно полная загрузка переносчика при полной отдаче в потребляющих тканях. Поэтому гиперболическая зависимость связывания от давления (как для миоглобина) не подходит (по данным рисунка и БфХ связывание при давлении 150 и 100 Торр близко к 100%, но для отдачи нужно очень низкое парциальное давление на уровне 1 Торр или даже менее – объяснение такого выбора кривой связывания миоглобина рассматривается ниже).
Уравнение Хилла Кривую насыщения гемоглобина (при характерном для теплокровных животных соотношении констант, обсуждение для холоднокровных – см. ниже) можно приближенно представить уравнением Хилла, т.е. зависимостью вида Y=pn/(Kn+pn), n = 2,8 Уравнение Хилла хорошо описывает реальную зависимость, если не рассматривать малые концентрации, где кривая насыщения линейна. Для человека и животных с близкой массой, значение К находится в диапазоне 3–4 кПа. Иллюстрация с подбором при значениях констант из БФХ (по исходной зависимости, уравнению Хилла + наиболее близкая к исходной гиперболическая кривая – как имитация возможностей приблизиться к наблюдаемому виду за счет одного связывания).
Физиологический конструктор По аналогии с молекулярным конструктором можно определить физиологический конструктор на основе удельных производительностей тканей и органов с учетом сопряжения. Производительные характеристики (биологические константы) и их зависимости (в том числе эмпирические) от условий (параметров) жизнедеятельности В простейшем варианте физиологический конструктор (и количественную физиологию) представляет набор биологических постоянных и зависимостей, объясняемых физико-химически (вариант: биохимией превращений как заданной), соображениями локальной целесообразности и/или эмпирически. К первому типу характеристик (объясняемых физико-химически), в частности, относятся: - энергетический эквивалент кислородного потребления (+ сама процедура измерения энергопотребления; обоснование – таблица 3), - характерное значение коэффициента обмена через поверхность для кислорода g = 0,33 л О2 м–2 <поверхности альвеол> ч–1 (его объясняют коэффициент диффузии кислорода в воде, размер клетки, механическая прочность покровов и кровеносного русла); - максимальный КПД превращения химической энергии в механическую – 20–25% (его объясняет стехиометрия биохимических превращений при получении АТФ и последующее использование АТФ в механизме мышечного сокращения).
Примеры второго типа характеристик (объясняемых некоторой узкой/локальной целесообразностью): - коэффициент обмена g через поверхность для кислорода одинаков для специализированных тканей, значение, рассчитанное по данным для млекопитающих из ФЖ (с.), можно использовать и шире (для дыхания через кожу и жабры – данные задачи про лягушку, там же, с.; расчет жабр); - содержание гемоглобина в крови (или сахаров во флоэмном соке) – см. ниже; - характерные концентрации интермедиатов биохимических превращений порядка 10–5 М (универсально как грубый выбор). Например, так можно обосновать характерную концентрацию субстратов биохимических превращений порядка 10–5 М (при том, что общее количество интермедиатов порядка сотни – для процессов относительно высокой интенсивности). С одной стороны, суммарную концентрацию всех веществ в клетке ограничивает осмотическое давление (25 атм на 1 М, поэтому существенным будет вклад в давление). С другой стороны, константа скорости лимитирующего этапа биохимических превращений имеет порядок kхим = 102-3 с-1 (Березин, Варфоломеев, «Биокинетика», с.) при диффузионной константе kD = 108-9 М-1 с-1. При таком соотношении констант концентрация субстрата S порядка 10–5 М уже не лимитирует, т.к. эффективная константа диффузионного этапа как произведение kD S на порядок меньше kхим. Есть также неспецифический эффект объемного вытеснения, определяемый почти исключительно молекулярной массой веществ (плотность органических веществ, отвечающая за их вклад в массу и объем раствора, близка к плотности воды, тогда на все вещества в ткани приходится около 200 г/л принимая, что 80% массы в ткани – это вода). Для высокомолекулярных составляющих предельное содержание в ткани можно оценить именно с этой точки зрения – см. ниже содержание миоглобина и субстратов дыхания.
Третий тип характеристик (эмпирические с некоторыми минимальными объяснениями – физико-химическими и др.) представляет набор зависимостей от массы для различных величин: - интенсивность основного обмена как потребление О2 для теплокровных I = 0,7 m 3/4 л О2/ч (m - масса в кг) и аналогично у холоднокровных I = 0,04 m 3/4 л О2/ч (если измерять его при 20оС). Наблюдаемые значения можно объяснять в связи с последовательным выбором активной формы поведения (см. ЕТЖ/далее); - затраты на перемещение вверх как mgv /КПД (хотя энергопотребление в целом включает и внутренние затраты, но по факту малы или в пределах оценки КПД), - площадь поверхности тела (м2) S (m) = 0,1 m 2/3 (общая тенденция подобия систематически не выполнена, но от этого фактора слабая зависимость + вырождение следует из-за взаимной компенсации различных изменений).
Связь производительных характеристик органов и тканей с молекулярными процессами (молекулярным конструктором). Пример: организация мускулатуры Наблюдаемая конкретная/детальная организация многих процессов вместе с некоторой естественной целесообразностью определяют их количественные характеристики. Например, интересно обсудить с этой точки зрения максимальную удельную силу сокращения мышц на единицу площади сечения мышцы – f = 40–50 Н/см2 (как следствие молекулярного механизма мышечного сокращения: затрата одной молекулы АТФ при характерном расстоянии дает силу одного элемента, а общее количество элементов на единицу площади сечения при наблюдаемой гексагональной структуре тонких и толстых филаментов составляет шт/см2 «Биофизика», с.) и затраты в связи со способностью мускулатуры к совершению сокращений Представление энергетических характеристик в характеристики напряжения: энергия связи = длина связи * сила (напряжение) ее разрыва (П) Бф, с. 408: связь молекулярных характеристик (N = 6,5 1012 мостиков на 1 см2 на половину саркомера длиной 1,1 мкм) с развиваемой силой (напряжение P0 = 300 кПа) дает силу f0 = P0/N = 4,6 10–12Н. При ориентировочной длине связи около l = 0,2 мкм и cosq = ½ получим произведение f0 l cosq = 4,6 10–22Дж, т.е. около 300 Дж/моль, что вроде бы маловато (в сравнении с ожидаемым значением порядка DG = 30-50 кДж/моль). развитие: оценка прочности различных молекулярных конструкций (например, фосфолипидного бислоя) и сравнение их с выдерживаемыми давлениями (например, в сосуде диаметром r натяжение T = P r, Бернулли/Лаплас по ФЖ, с. 166) Важное следствие молекулярного механизма сокращения (когда несогласованно образуются и разрываются мостики между тонкими и толстыми филаментами, альтернативный вариант одновременного образование и разрыва потребовал бы несоизмеримо более сложной организации и невозможен в силу непреодолимого активационного барьера): поддержание силовой нагрузки требует тем больших затрат, чем потенциально более быстрой является мускулатура. При такой организации затраты линейно связаны с потенциально развиваемой силой. В частности, поддержание статической нагрузки (когда совершаемая мускулатурой механическая работа равна нулю) требует работы и энергопотребления. В этом смысле экономия затрат состоит в том, чтобы поддерживать статическую или близкую к ней нагрузку за счет медленной мускулатуры – в пределе очень медленной мускулатуры (и по факту, например, для замыкающей мышцы моллюсков) работа близка к нулю в статическом режиме. Простые возможности расчетов на основе физиологического конструктора: –негативные оценки Использование набора характеристик физиологического конструктора в простейшем варианте – «Задачи по количественной физиологии»: необходимость в органах дыхания в зависимости от размера теплокровных и холоднокровных организмов; мелкие организмы на единицу массы сильнее крупных Задача о дыхании на высоте. Известно, что интенсивность потребления кислорода человеком при интенсивной мышечной нагрузке может возрастать на порядок, а у спортсменов – в 20–25 раз. При этом потребление кислорода в мышцах может возрастать даже в 100 раз [Шмидт-Ниельсен, 1982, с. 173]. Почему же тогда человек, вдыхая воздух, способен выжить лишь несколько часов на высоте 8600 м (где атмосферное давление составляет 250 мм рт.ст., т.е. уменьшается по сравнению с уровнем моря примерно в 3 раза). Как удается летать на такой высоте птицам (при неизбежно сопровождающей полет интенсивной мышечной нагрузке)? Как при уменьшении парциального давления кислорода изменяется скорость отдельных этапов поступления кислорода в организм? –расчет органов и систем Наиболее интересен анализ ситуаций, в которых наиболее явно выражены проблемы некоторого рода, как основа для расчета соответствующих органов. Примеры заданий к семинарам (в отдельном тексте): жираф – расчет органов дыхания (у которого очень длинная шея, а значит большой объем «мертвого пространтства); сравнение альтернатив организации – дыхание воздухом как в воде и наоборот.
Введение в количественную биологию Васильев Алексей Артёмович при участии Окштейна Игоря Леонидовича (семинары в рамках лекционно-семинарского курса «Основы биологии») Часть 2 (весенний семестр)
|
||||
|
|
Последнее изменение этой страницы: 2016-04-08; просмотров: 163; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.148.145.219 (0.009 с.) |




 …
… 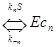
 ,
,


