
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Т. П. Вавилова, Ю. Г. Гаверова, О. Н. Гева, Н. И. ДеркачеваСтр 1 из 9Следующая ⇒
МЕДИКО-СТОМАТОЛОГИЧЕСКИЙ УНИВЕРСИТЕТ ИМЕНИ А.И.ЕВДОКИМОВА» Т.П. Вавилова, Ю.Г. Гаверова, О.Н. Гева, Н.И. Деркачева ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ МЕТАБОЛИЧЕСКИХ ПРОЦЕССОВ Учебно-методическое пособие МОСКВА, 2018 УДК: 577.7 (075.8) ББК 28.707.2я73 Б 63 Рецензенты: Зав. кафедрой общей и биоорганической химии МГМСУ, д.х.н, профессор Прокопов А.А. Ведущий научный сотрудник НИИ Физико-Химической биологии им. А.Н. Белозерского МГУ им. М.В. Ломоносова, д.б.н., профессор Соловьев А.Г.
Гормональная регуляция метаболических процессов. Учебно-методическое пособие: д.м.н., профессор Т.П. Вавилова, к.б.н., доцент Ю.Г. Гаверова, О.Н. Гева, к.б.н., доцент Н.И. Деркачева. Изд. МГМСУ, кафедра биохимии, зав. каф. д.м.н, профессор Т.П. Вавилова, 2018, стр.48
Учебно-методическое пособие «Гормональная регуляция метаболических процессов» подготовлено в соответствии с Федеральным государственным образовательным стандартом третьего поколения. Представленный материал охватывает широкий спектр вопросов, связанных с передачей сигнала и влиянием на обменные процессы всех основных гормонов организма человека. Данное пособие может использоваться студентами стоматологического и лечебного факультетов при подготовке к текущим занятиям и к экзаменам по дисциплинам «Биохимия» и «Биохимия полости рта».
Утверждено на заседании Учебно-методического совета ФГБОУ ВО МГМСУ им. А.И. Евдокимова Минздрава России протокол № 4 от 25.04.2018
© Т.П. Вавилова, Ю.Г. Гаверова, О.Н. Гева, Н.И. Деркачева, 2018 © МГМСУ имени А.И. Евдокимова Оглавление
Механизмы передачи гормонального сигнала
Рецептор к гормону может находиться внутри клетки или на плазматической мембране. Локализация рецептора зависит от химической природы гормона. Соматотропный гормон Гормон роста (соматотропин) секретируется аденогипофизом в количествах в десятки раз превышающих секрецию других тропных гормонов. Секреция соматотропина происходит с интервалами 20-30 минут в течение всего дня, однако наибольшее его количество выделяется во сне. Основным стимулирующим секрецию фактором является соматолиберин, а тормозящим – соматостатин, однако секреция может существенно повышаться под действием физических упражнений, стресса, голодания и белковой диеты. Основная функция соматотропина заключается в регуляции процессов, связанных с ростом и развитием организма. Под влиянием этого гормона происходит рост различных органов и тканей, в том числе костей, мышц и внутренних органов. Рецепторы гормона роста находятся фактически во всех тканях организма. Взаимодействие соматотропина с рецептором активирует тирозинкиназную и инозитолфосфатную системы передачи сигнала. Действие гормона роста опосредуется с помощью особых биологически активных молекул соматомединов (инсулиноподобных факторов роста или ИФР), которые образуются в печени в ответ на связывание соматотропина с рецепторами. Наиболее выраженным эффектом обладает ИФР-1, повышение секреции которого по принципу отрицательной обратной связи тормозит секрецию соматотропина. Инсулиноподобные факторы роста, как и инсулин, передают сигнал через тирозинкиназный рецептор и вызывают мощные ростовые эффекты.
Гормон роста активирует синтез белка в мышечной, хрящевой, костной и других тканях организма. Под его воздействием усиливается транспорт аминокислот в клетки, синтез РНК и ДНК, активируются клеточные деления. Первичные кратковременные эффекты гормона роста на обмен веществ похожи на действие инсулина: усиление синтеза липидов в жировой ткани, снижение уровня глюкозы в крови. Более поздние долговременные влияния на обмен углеводов и жиров противоположны действию инсулина. Происходит активация тканевого липолиза, что приводит к повышению уровня жирных кислот в плазме крови. Активируется β-окисление жирных кислот, полученная при этом энергия используется для поддержания анаболических процессов, сопровождающих рост. Под действием соматотропина повышается уровень глюкозы в крови, как за счет уменьшения потребления ее мышцами и жировой тканью, так и за счет активации глюконеогенеза. При длительном введении соматотропина в терапевтических целях или у спортсменов может развиваться сахарный диабет. Соматотропин активирует синтез сульфатированных гликозаминогликанов, способствуя формированию органического матрикса хрящевой и костной тканей. Повышая продукцию ИФР-1, соматотропин оказывает влияние на минеральный обмен, способствует минерализации костной ткани, вызывает задержку ионов Na, K и Cl в организме. Недостаточная секреция гормона роста у детей приводит к развитию карликовости или нанизма. Наиболее частая причина карликовости – мутация в гене гормона роста или его рецептора. Избыточная секреция гормона роста наблюдается при аденоме гипофиза. У половины больных с гиперсекрецией гормона роста наблюдается мутация в гене α-субъединицы G-белка в клетках гипофиза, продуцирующих гормон роста. При этой мутации у α-субъединицы снижается ГТФ-азная активность и при передаче сигнала от соматолиберина происходит избыточная активация аденилатциклазы, что приводит к избыточной секреции соматотропина. При гиперсекреции соматотропина у детей развивается гигантизм, а у взрослых – акромегалия. При гигантизме увеличение частей тела и внутренних органов происходит относительно пропорционально. При акромегалии под действием избытка соматотропина происходит непропорциональное увеличение надбровных дуг, скуловых костей, стоп и кистей, разрастание мягких тканей лица. Кроме того, может происходить значительное увеличение внутренних органов. Повышенная смертность больных акромегалией наблюдается из-за болезней сердца и легких, а также вследствие поражения сосудов головного мозга. Пролактин Гормон пролактин выделяется определенными клетками аденогипофиза и похож по строению и механизму передачи сигнала на гормон роста. Рецепторы пролактина имеются в печени, почках, надпочечниках, половых железах, матке и других тканях. Секреция пролактина усиливается под действием тиреолиберина, серотонина, окситоцина, ацетилхолина, а также эстрогенов. Дофамин оказывает тормозящее действие на выделение пролактина, кроме того секрецию пролактина тормозит прогестерон.
Основная функция пролактина у женщин – это стимуляция лактации. В молочной железе под действием пролактина усиливается синтез белков и липидов. У мужчин пролактин регулирует секрецию тестостерона. Пролактин оказывает воздействие на работу почек, способствуя реабсорбции воды, ионов Na+ и К+. Кроме того, пролактин повышает иммунный ответ организма. Тиреотропин Тиреотропин (тиреотропный гормон, ТТГ) вырабатывается клетками передней доли гипофиза и действует на клетки щитовидной железы по аденилатциклазному механизму. Он стимулирует все этапы синтеза иодтиронинов. Непрерывное действие ТТГ в течение нескольких дней вызывает разрастание ткани щитовидной железы. Возрастает количество фолликулярных клеток, что приводит к усилению секреции тиреоидных гормонов. Синтез и секреция ТТГ регулируются по принципу обратной связи и зависят от количества тиреолиберина и иодтиронинов. Гонадотропины К гонадотропным гормонам относятся фолликулостимулирующий гормон (ФСГ, фоллитропин) и лютеинизирующий (ЛГ, лютропин). Они реализуют свой сигнал по аденилатциклазному механизму. У мужчин эти гормоны секретируются в пульсирующем режиме. В семенниках ЛГ стимулирует синтез и секрецию тестостерона, а ФСГ усиливает синтез рецептора тестостерона, трансферрина, эстрогенов, активатора плазминогена. В женском организме все несколько сложнее, Секреция гонадотропинов подчиняется менструальному циклу. ФСГ в первую фазу стимулирует в яичниках секрецию эстрогенов. Повышение концентрации эстрогенов стимулирует секрецию ЛГ, который стимулирует овуляцию и развитие желтого тела. Желтое тело производит прогестерон и дополнительное количество эстрогенов, что приводит к угнетению секреции ФСГ и ЛГ. Последующее снижение концентрации эстрогенов и прогестерона, в случае если беременность не наступила, возвращает организм женщины в первую фазу цикла. При наступлении беременности функцию синтеза и секреции эстрогенов и прогестерона берет на себя плацента, и в этом случае секреция гонадотропинов остается на низком уровне. Вазопрессин (АДГ) Рецепторы к вазопрессину находятся в миоцитах кровеносных сосудов и в клетках почечных канальцев. В собирательных трубочках и дистальных канальцах почек вазопрессин взаимодействует с V2-рецепторами, что обеспечивает передачу сигнала по аденилатциклазному механизму. В результате происходит активация белков, которые запускают транскрипцию генов белков аквапоринов-2, обеспечивающих реабсорцию воды в почечных канальцах. Снижение диуреза и является основным физиологическим эффектом гормона, поэтому он называется антидиуретическим. В кровеносных сосудах вазопрессин взаимодействует с V1-рецепторами и передает сигнал через инозитолфосфатную систему, что приводит к сокращению просвета кровеносного сосуда и повышению артериального давления. Благодаря способности повышать тонус кровеносных сосудов гормон и называется вазопрессином. Взаимодействие гормона с V1-рецепторами происходит только при его повышенной концентрации в крови.
Секреция вазопрессина повышается при возрастании осмотического давления плазмы крови и внеклеточной жидкости: при обильном потоотделении, жажде, стрессе, кровопотерях, а также при потреблении большого количества соли. Наиболее высокая базовая секреция гормона наблюдается в ночное время суток. Вазопрессин оказывает положительное влияние на память и внимание. Снижение секреции АДГ приводит к нарушению реабсорбции воды из первичной мочи и развитию несахарного диабета (термин «диабет» происходитотгреческого слова diabetes (от diabaino - прохожу) и является общим названием группы болезней, характеризующихся избыточным выделением мочи). Общий суточный диурез может достигать 20 литров в сутки, плотность мочи снижается. Несахарный диабет приводит к дегидротации организма. Причинами несахарного диабета могут быть мутации в генах вазопрессина и V2-рецептора, а также черепно-мозговые травмы, последствия инсультов головного мозга, опухоли. Окситоцин Мембранные рецепторы окситоцина найдены в матке и молочной железе. Передача сигнала происходит с участием инозитолфосфатной системы, при этом ведущее значение имеет изменение концентрации ионов Са2+в клетке. В лактирующей молочной железе под действием окситоцина происходит сокращение миоэпителиальных клеток выводных протоков и молоко, выработанное под действием пролактина выделяется из груди. Окситоцин стимулирует сокращение гладкой мускулатуры матки при родах и используется для стимуляции родовой деятельности. Окситоцин негативно влияет на память и внимание, что, вероятно, снижает стрессовое воздействие родовых болей на организм роженицы.
Гормоны щитовидной железы В щитовидной железе синтезируется два типа гормонов, отличающихся по химической природе и метаболическим эффектам. Йодтиронины, или тиреоидные гормоны, являются йодированными производными аминокислоты тирозина. Их действие преимущественно направлено на регуляцию энергетического обмена веществ, а также роста и развития организма. Гормон кальцитонин имеет белковую природу, он участвует в регуляции фосфорно-кальциевого обмена и ремоделировании костной ткани.
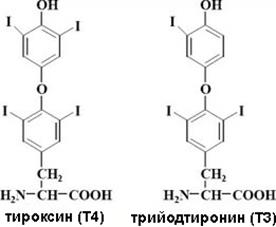
Йодтиронины К йодтиронинам относятся тироксин или тетрайодтиронин, содержащий 4 атома йода (Т4) и трийодтиронин (Т3) (рис.10). Активность Т3 в несколько раз превышает активность Т4, и хотя в плазме крови Т3 содержится в 50 раз меньше, чем Т4, но именно Т3 обеспечивает эффекты йодтиронинов. В настоящее время Т4 рассматривается в качестве прогормона, так как в периферических тканях происходит его дейодировании под действием 5'-дейодиназы типа I с образованием Т3. В активном центре дейодиназ йодтиронинов находится остаток селенцистеина.
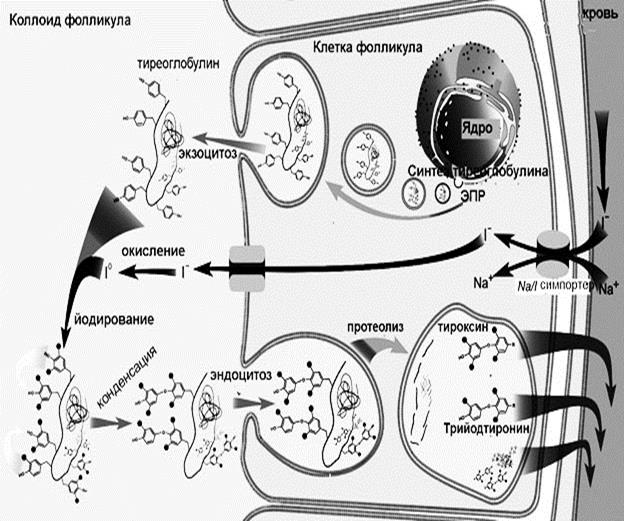
Рис. 10. Структурные формулы йодтиронинов. В сыворотке крови большая часть тиреоидных гормонов связано с альбуминами и глобулинами, и только 0,03% Т4 и 0,3% Т3 находится в свободном состоянии, но именно свободные гормоны обладают физиологической активностью. В клетках щитовидной железы образование тиреоидных гормонов начинается с синтеза белка тиреоглобулина, богатого остатками тирозина (рис.11). Тиреоглобулин выводится в просвет фолликулов щитовидной железы путем экзоцитоза. В коллоид фолликула также поступает йод в форме йодида, который окисляется под действием йодидпероксидазы, что необходимо для осуществления йодирования остатков тирозина в составе тиреоглобулина. Йодирование приводит к образованию монойодтирозинов и дийодтирозинов, которые вступают в реакцию конденсации друг с другом. Конденсация двух молекул дийодтирозина приводит к образованию тироксина, а при конденсации дийодтирозина с монойодтирозином образуется трийодтиронин. Конденсация происходит в составе тиреоглобулина, который затем путем эндоцитоза попадает обратно в клетку фолликула, где подвергается протеолизу во вторичных лизосомах. В ходе протеолиза происходит высвобождение молекул тироксина и трийодтиронина, которые затем поступают в кровь.
Рис.11. Схема синтез йодтиронинов. Рецепторы тиреоидных гормонов находятся в ядре клеток, гормон-рецепторные комплексы активируют транскрипцию определенных генов, в результате происходит увеличение скорости синтеза соответствующих белков. Физиологическое действие йодтиронинов прежде всего направлено на регуляцию роста и развития организма. Трийодтиронин усиливает экспрессию гена гормона роста. Йодтиронины стимулируют пролиферацию, дифференцировку и рост клеток различных тканей. Основные метаболические эффекты тиреодиных гормонов направлены на активацию энергетического обмена. Йодтиронины индуцируют синтез ферментов общих путей катаболизма, в первую очередь белковых комплексов дыхательной цепи. В результате происходит увеличение потребления кислорода и продукции АТФ. С другой стороны йодтиронины индуцируют синтез мембранных АТФ-аз, в том числе Nа+,К+-АТФ-азы, что способствует потреблению образованного АТФ. Йодтиронины обеспечивают повышение теплопродукции при охлаждении организма. Этот эффект реализуется за счет усиления тканевого липолиза, продукты которого, свободные жирные кислоты, выполняют роль разобщителей в дыхательной цепи. Увеличение концентрации жирных кислот создает условия, в которых большая часть энергии трансмембранного электрохимического потенциала дыхательной цепи митохондрий рассеивается в виде тепла. В печени йодтиронины активируют глюконеогенез, мобилизацию гликогена, синтез холестерола и желчных кислот. В мышцах под действием Т3 возрастает потребление глюкозы и активируется синтез белка, что приводит к увеличению мышечной массы. Важным эффектом йодтиронинов является усиление действия катехоламинов. Кальцитонин Кальцитонин - пептид, состоящий их 32 аминокислот, его синтез осуществляется не только в парафолликулярных клетках щитовидной железы, но также и в паращитовидных железах. Этот гормон передает сигнал в клетку через аденилатциклазную систему. Секреция кальцитонина усиливается в ответ на повышение концентрации ионов кальция в плазме крови. По своим эффектам кальцитонин является антогонистом паратгормона (см. раздел 5). Кальцитонин снижает реабсорбцию Са2+ в почечных канальцах и тем самым способствует экскреции этого иона с мочой. Рецепторы к кальцитонину находятся на остеокластах, его действие тормозит их созревание и активность, что приводит к снижению скорости резорбции костной ткани.
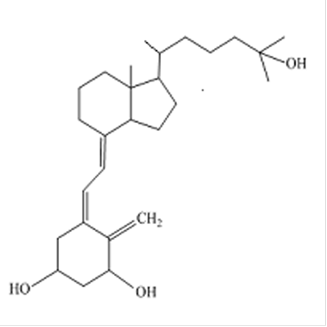
Гормон паращитовидных желез Паращитовидные железы секретируют паратгормон, который представляет собой белок, состоящий из 84 аминокислот. В целом эффект этого гормона заключается в повышении уровня ионов кальция и снижение уровня фосфатов в плазме крови. Паратгормон передает сигнал по аденилатциклазному механизму. Секреция гормона повышается при снижении уровня ионов кальция в плазме крови. Паратгормон является антогонистом кальцитонина. Рецепторы к парагормону находятся в почках и в костной ткани. В костной ткани рецепторы к паратгормону находятся на остеобластах и остеоцитах. Связывание паратгормона с этими рецепторами стимулирует секрецию факторов, вызывающих образование и активацию остеокластов. Это приводит к усилению резорбции костной ткани. Разрушение минерализованного межклеточного матрикса костной ткани приводит к выделению ионов кальция и фосфатов во внеклеточную жидкость. В почках паратгормон усиливает реабсорбцию ионов кальция и экскрецию фосфатов, что приводит к повышению уровня кальция и снижению уровня фосфатов в плазме крови. Кроме того, паратгормон активирует синтез кальцитриола. Кальцитриол представляет собой 1,25-дигидроксихолекальциферол, активную форму витамина D3 (холекальциферола) (рис.12).
Рис. 12. Структурная формула кальцитриола Синтез холекальциферола из 7-дегидрохолестерола происходит в коже под действием ультрафиолетового излучения. Кальцитриол образуется путем гидроксилирования холекальциферола в печени по 25-ому положению и в почках по 1-ому положению. Паратгормон активирует 1α-гидроксилазу почек и тем самым стимулирует образование кальцитриола. В энтероцитах кальцитриол, передавая сигнал по цитозольно-ядерному механизму, индуцирует синтез мембранных белков, осуществляющих транспорт ионов кальция из просвета кишечника в клетку. Таким образом, кальцитриол усиливает всасывание кальция в кишечнике, что приводит к повышению его концентрации в плазме крови. В клетках почечных канальцев кальцитриол индуцирует синтез белков, участвующих в реабсорбции ионов кальция и фосфатов из первичной мочи. В результате усиления всасывания и реабсорбции повышается уровень ионов кальция и фосфатов в плазме крови. В остеобластах и остеоцитах кальцитриол индуцирует синтез Са2+-связывающего белка остеокальцина. Этот белок, с одной стороны участвует в процессе минерализации, а с другой, способствует прикреплению остеокластов к участку резорбции. В результате суммарный эффект повышения уровня кальция в плазме крови под действием паратгормона происходит за счет усиления его реабсорции в почках и поступления из слизистой оболочки кишечника, а также выделения из костной ткани.
Таблица 1. Влияние инсулина на метаболические процессы
Увеличение скорости | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Гликолиз | Глюкокиназа | Индукция синтеза | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Фосфофруктокиназа | Активация | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Пируваткиназа | Активация, индукция синтеза | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Окислительное декарбоксилирование пирувата | Пируватдегидрогеназный комплекс | Активация | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Синтез гликогена | Гликогенсинтаза | Активация | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Пентозофосфатный путь | Глюкозо-6-фосфатдегидрогеназа | Индукция синтеза | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Синтез жирных кислот | Ацетил-КоА-карбоксилаза | Активация, индукция синтеза | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Пальмитатсинтаза | Индукция синтеза | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Синтез холестерола | ГМГ-КоА-редуктаза | Активация | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Снижение скорости | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Глюконеогенез | Фосфоенолпируват- карбоксикиназа | Репрессия синтеза | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Глюкозо-6-фосфатаза | Ингибирование | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Распад гликогена | Гликогенфосфорилаза | Ингибирование | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Тканевой липолиз | ТАГ-липаза | Ингибирование | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Гормоны надпочечников
Надпочечники – это парные эндокринные органы, расположенные на верхних полюсах почек. По своему строению и секреторным функциям надпочечники неоднородны (рис. 13). Наружная часть надпочечников носит название корковое вещество или кора, в нем происходит синтез стероидных гормонов (кортикостероидов). Мозговое вещество надпочечников представляет собой видоизмененный узел симпатической нервной системы, в нем происходит синтез катехоламинов.
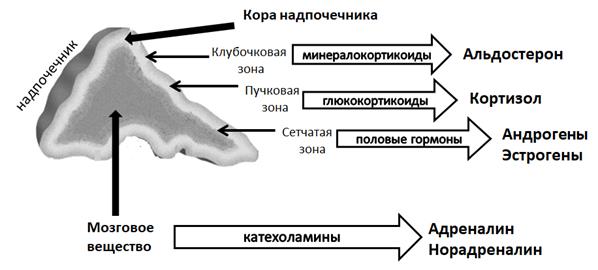
Рис.13. Гормоны надпочечников
Гормоны коры надпочечников
В коре надпочечников выделяются три зоны, каждая из которых продуцирует определенный вид кортикостероидов (рис.13). Кортикостероиды являются производными холестерола и передают сигнал по цитозольно-ядерному механизму, оказывая воздействие на транскрипцию генов. Самый наружный слой коры, клубочковая зона, вырабатывает минералокортикоиды, которые регулируют водно-солевой обмен и способствуют сохранению ионов Na+ в плазме крови и выведению ионов К+. Наиболее активным минералокортикоидом является альдостерон. Пучковая зона осуществляет синтез глюкокортикоидов, участвующих в регуляции обмена углеводов, липидов, белков, и, соответственно, связанных с энергетическим обменом. Основным глюкокортикоидом является кортизол. Самый глубокий слой коры, сетчатая зона, участвует в синтезе половых гормонов, преимущественно андрогенов, из которых в небольших количествах образуются эстрогены. Половые гормоны коры надпочечников играют важную роль до полового созревания, после полового созревания основную роль в их синтезе выполняют половые железы. Структурные формулы гормонов коры надпочечников представлены на рисунке 15.
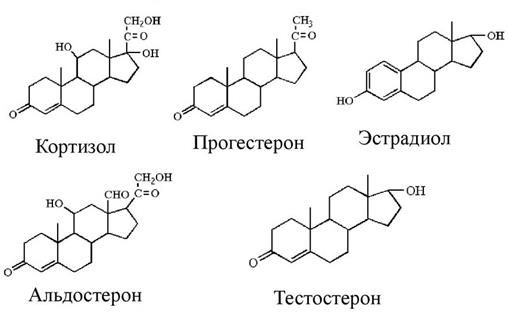
Рис. 15. Структурные формулы гормонов коры надпочечников
Глюкокортикоиды
В целом действие глюкокортикоидов направлено на обеспечение функционирование организма в условиях длительного стрессового воздействия.
Основным эффектом кортизола на обмен углеводов является увеличение скорости глюконеогенеза за счет индукции синтеза ключевого фермента этого процесса фосфоенолпируваткарбоксикиназы. В непеченочных тканях (особенно в мышечной) под действием кортизола происходит катаболизм белков, а освобождающиеся при этом аминокислоты поступают в печень, дезаминируются и используются в процессе глюконеогенеза. В печени кортизол индуцирует синтез аланинаминотрансферазы (АЛТ) и других ферментов распада аминокислот. Таким образом, глюкокортикоиды способствуют превращению аминокислот в глюкозу и повышению уровня глюкозы в крови, что обеспечивает глюкозой мышцы, мозг и другие периферические ткани при стрессе. Влияние глюкокортикоидов неодинаково в разных тканях и органах. В печени их действие приводит к анаболическим эффектам: происходит активация синтеза белков и нуклеиновых кислот. Кортизол в избытке усиливает липолиз в конечностях и способствует отложению жира в районе туловища и лица.
Повышенные концентрации глюкокортикоидов тормозят иммунные реакции, ослабляют воспалительные процессы и аллергические реакции. Данные эффекты обуславливают применение этих гормонов в качестве лекарственных средств. В высокой концентрации глюкокортикоиды тормозят деление фибробластов, ингибируют синтез коллагена и других белков межклеточного матрикса.
Минералокортикоиды
Наиболее активным минералокортикоидом является альдостерон, основными органами-мишенями которого являются почки. Действие альдостерона направлено на уменьшение выведения ионов Na+ и увеличение выведения ионов К+ в почечных канальцах. Аналогичное действие альдостерон оказывает на работу слюнных и потовых желез.
Синтез и секреция альдостерона повышается при снижении концентрации ионов Na+ и увеличении ионов К+ в плазме крови. Секреция альдостерона находится по контролем гипоталамо-гипофизарной системы и повышается под действием АКТГ. Наибольшее влияние на секрецию альдостерона оказывает ренин-ангиотензиновая система.
Ренин – это протеаза, которая образуется в юкстагломерулярных клетках артериол почечных клубочков. Секреция ренина возрастает при снижении артериального давления вследствие кровопотери, рвоты или других факторов, вызывающих обезвоживание организма. Под действием ренина происходит частичный протеолиз белка ангиотензиногена, который присутствует в плазме крови в составе фракции α2-глобулинов, и в результате образуется состоящий из 10 аминокислот неактивный пептид, ангиотензин I. Ангиотензин I под действием ангиотензин-превращающего фермента, вырабатывающегося преимущественно в эндотелии сосудов и в легких, превращается в активный ангиотензин II, состоящий из 8 аминокислот. Ангиотензин II, передавая сигнал через аденилатцилазную систему, непосредственно стимулирует выработку альдостерона в коре надпочечников. Активируя инозитолфосфатную систему, ангиотензин II вызывает сокращение гладких мышц кровеносных сосудов, что приводит к повышению артериального давления. Под действием аминопептидаз ангиотензин II частично превращается в ангиотензин III, снижающий активность ангиотензина II.
Альдостерон индуцирует синтез белков-транспортеров Na+ и К+, что способствует увеличению реабсорбции натрия из первичной мочи в клетки почечного канальца и выведению калия в первичную мочу. Под действием альдостерона происходит индукция синтеза Na+, К+-АТФ-азы, что приводит к поступлению ионов Na+ из клеток в кровь, а ионов К+ - в клетки почечных канальцев. И, наконец, индукция альдостероном синтеза ключевого фермента цикла Кребса цитратсинтазы позволяет увеличить образование АТФ в дыхательной цепи для обеспечения работы Na+, К+-АТФ-азы.
Таким образом, суммарный эффект альдостерона приводит к увеличению концентрации ионов Na+ в плазме крови, и соответственно, к задержке воды и увеличению артериального давления. Повышение концентрации ионов Na+ в плазме крови служит сигналом для секреции антидиуретического гормона в задней доле гипофиза, что также способствует сохранению воды в организме и подъему артериального давления. Схема работы системы ренин-ангиотензин-альдостерон представлена на рисунке 16.
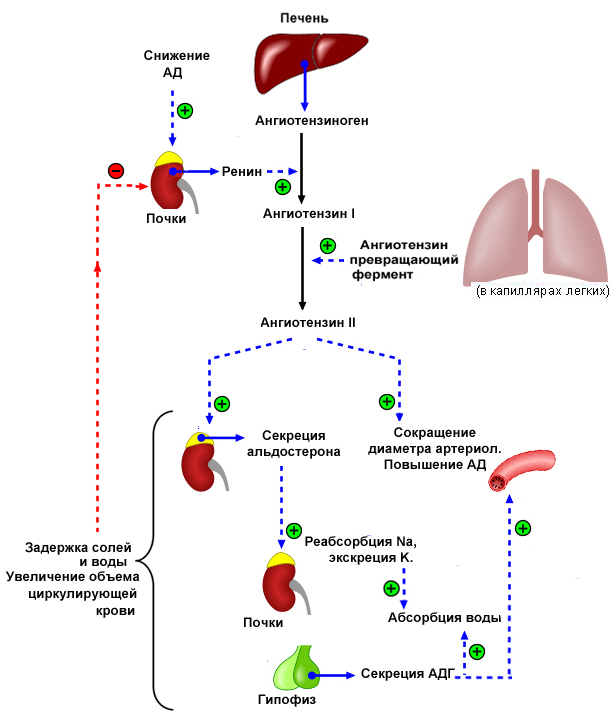
Рис. 16. Ренин-ангиотензин-альдостероновая система
Антагонистом альдостерона и ангиотензина II является предсердный натрийуретический фактор, имеющий белковую природу и образующийся преимущественно в кардиомиоцитах предсердий. Мембранный рецептор этого гормона обладает гуанилатциклазной активностью. Секреция натрийуретического фактора повышается при возрастании артериального давления, увеличении частоты сердечных сокращений, а также под действием адреналина и глюкокортикоидов.
Рецепторы к натрийуретическому фактору находятся в почках и периферических артериях. В почках он стимулирует увеличение общего
кровотока и выведения ионов натрия. Под действием натрийуретического фактора снижается тонус гладких мышц кровеносных сосудов. Таким образом, суммарный эффект этого гормона выражается в снижении артериального давления.
Гормоны половых желез
Женские и мужские половые гормоны являются производными холестерола и передают сигнал по цитозольно-ядерному механизму. У взрослого основное количество половых гормонов синтезируется в половых железах, однако невысокий уровень синтеза имеет место и в сетчатой зоне коры надпочечников. Секреция половых гормонов контролируется гипоталамо-гипофизарной системой. Основной функцией этих гормонов является регуляция репродуктивных функций организма, а также поддержание обмена веществ в наиболее благоприятном состоянии для осуществления этих функций.
Мужские половые гормоны
В семенниках происходит синтез мужских половых гормонов, основным из которых является тестостерон (рис. 17). Синтез тестостерона стимулирует лютеинизирующий гормон передней доли гипофиза.
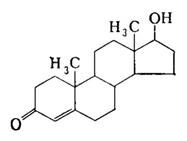
Рис. 17. Структурная формула тестостерона
Тестостерон стимулирует развитие семенников в эмбриогенезе и регулирует сперматогенез у взрослого организма, а также влияет на формирование вторичных мужских половых признаков. Под действием андрогенов происходит утолщение голосовых связок, что приводит к снижению тембра голоса. Андрогены стимулируют секрецию сальных желез и вызывают изменение структуры волос
Андрогены обладают выраженным анаболическим эффектом, они усиливают синтез белка, в первую очередь, в костной, хрящевой и мышечной тканях. Эти гормоны ускоряют тканевой липолиз, активируют синтез нуклеотидов и деление клеток.
Совместно с гормоном роста андрогены стимулируют рост костей с последующим закрытием зон роста под действием повышенного уровня секреции. Мужские половые гормоны оказывают положительное влияние на рост мышц и увеличивают продукцию эритроцитов.
Женские половые гормоны
В яичниках образуются женские половые гормоны, эстрогены и прогестины. Наиболее активными среди них являются 17β-эстрадиол и прогестерон (рис. 18). Эстрогены синтезируются из андрогенов с участием реакций микросомального окисления. Этот процесс стимулирует фолликулстимулирующий гормон (ФСГ), секретируемый в передней доле гипофиза. Существенная часть эстрогенов синтезируется в желтом теле, коре надпочечников, печени, жировой ткани и коже, а при беременности – в плаценте.

Рис. 18. Структурные формулы женских половых гормонов
Эстрогены стимулируют развитие тканей, участвующих в размножении, определяют развитие женских вторичных половых признаков, индуцируют синтез рецептора прогестина и увеличивают чувствительность матки к действию окситоцина перед родами.
Эстрогены обладают выраженными анаболическими эффектами. Они усиливают синтез белков, стимулируют липогенез и синтез холестерола. Под действием эстрогенов в крови повышается уровень ЛВП, что способствует нормализации транспорта холестерола. Увеличение под действием эстрогенов продукции NO клетками эндотелия приводит к расширению кровеносных сосудов. Эстрогены индуцируют синтез факторов свертывания крови.
Прогестерон образуется преимущественно в желтом теле, а после 6-ой недели беременности – в плаценте. Этот гормон регулирует протекание менструального цикла, готовит матку к беременности и родам, обеспечивает нормальное протекание беременности.
Задания для контроля знаний.
1. Дополните следующие утверждения:
Гормон может воздействовать на клетку, если в ней присутствует специфический _____________.
Клетки, содержащие рецептор к гормону, называются _______________.
Локализация рецептора в клетке зависит от _______________ _________ ________________
При передаче сигнала по цитозольно-ядерному механизму гормон-рецепторный комплекс влияет на ___________ _____________.
При передаче сигнала по аденилатциклазному механизму a-субъединица в комплексе с ГТФ активирует фермент _______________.
При передаче сигнала по инозитолфосфатному механизму a-субъединица в комплексе с ГТФ активирует фермент _______________________.
Гормоны гипоталамо-гипофизарной системы имеют химическую природу _____________.
Координация нервной и эндокринной регуляции в организме человека осуществляется за счет работы системы ____________________________.
Рецепторы к либеринам и статинам гипоталамуса находятся в мембранах клеток ________________________________.
Большинство гормонов гипоталамо-гипофизарной системы передают сигнал по ______________________ и _____________________ механизмам.
Вазопрессин и окситоцин синтезируются в _______________, а поступают в кровяное русло в ____________ __________ ________________.
При избыточной секреции гормона роста у взрослых развивается _______________________.
В клетках щитовидной железы синтез йодтиронинов осуществляется в составе белка _____________.
Основной механизм передачи сигнала для йодтиронинов ____________________________.
Кальцитонин передает сигнал по ______________________ механизму.
Паратгормон передает сигнал по ______________________ механизму.
Кальцитриол – это активная форма витамина _____________.
Кальцитриол передает сигнал по механизму _______________________.
Инсулин передает сигнал по механизму _________________________
Инсулин по химической природе является _______________________.
Снижение секреции инсулина приводит к развитию ___________________.
Наиболее активным минералокортикоидом является __________________.
Наиболее активным глюкокортикоидом является ____________________.
Ренин – это ______________, которая вырабатывается в _______________________________________________________.
МЕДИКО-СТОМАТОЛОГИЧЕСКИЙ УНИВЕРСИТЕТ
ИМЕНИ А.И.ЕВДОКИМОВА»
Т.П. Вавилова, Ю.Г. Гаверова, О.Н. Гева, Н.И. Деркачева



