Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Очень быстрые эффекты (секунды)Содержание книги
Поиск на нашем сайте
1. Гиперполяризация мембран чувствительных к инсулину клеток;
2. Активация na+/h+-обменника, что выхывает выход ионов н+, вход в клетку ио-
нов na+;
3. Активация na+/к+-обменника, что выхывает выход ионов na+, вход в клетку ио-нов k+;
4. Угнетение ca2+-насоса, приводит к задержке ионов са2+ в клетке;
5. Стимуляция транспорта глюкозы в клетку – появление на мембране перенос-чиков глюкозы;
Быстрые эффекты (минуты)
1. Стимулирование протеинфосфатаз приводит к активации гликогенсинтазы,
пируватдегидрогеназы, ГМГ-SКоА-редуктазы, ацетил-S-КоА-карбоксилазы;
2. Увеличивает липогенез за счет создания благоприятного "биохимического
климата":
• активирует глюкозо-6-фосфатдегидрогеназу и, что вызывает наработку НАДФН, • глюкокиназу, что ведет к синтезу ацетил-SКоА,
• ацетил-S-КоА-карбоксилазу и синтазу жирных кислот, повышая синтез жир-ных кислот. 3. Активирует цАМФ-фосфодиэстеразу, снижая тем самым ее активирующее
влияние на протеинкиназу А и реакции катаболизма.
Медленные эффекты (минуты-часы)
1. Активация синтеза глюкокиназы, АТФ-цитрат-лиазы, ацетил-S-КоА-карбо-
ксилазы, синтазы жирных кислот, пируваткиназы, люкозо-6-фосфатдегидрогеназы, цитолозольной малатдегидрогеназы.; 2. Увеличение синтеза тРНК для увеличения скорости транскрипции. Однако на-работку мРНК антагонистических ферментов снижает (например, для ФЕП-
карбоксикиназы);
3. Повышает фосфорилирование по серину рибосомального белка S6, что сти-мулирует синтез белка.
Очень медленные эффекты (часы-сутки)
1. Повышает синтез соматомедина, зависимый от гормона роста;
2. Увеличивает рост и пролиферацию клеток, действуя при этом синергично с
соматомедином;
3. Стимулирует тирозиновые киназы., вызывает переход клетки из G1 в S-фазу клеточного цикла.
П ОСЛЕДСТВИЯ ДЕФИЦИТА ИНСУЛИНА
Быстрые последствия
1. Гипергликемия –так как отсутствует влияние инсулина и превалирует влия-
ние глюкагона, адреналина, кортизола, гормона роста.
2. Глюкозурия –почечный порог для глюкозы,т.е.концентрация глюкозы в кро-
ви при которой она появляется в моче, примерно равен 10,0 ммоль/л. В норме в мо-
че уровень глюкозы 0,8 ммоль/л и до 2,78 ммоль/сут, в других единицах около 0,5
г/сут, при СД количество теряемой глюкозы составляет до 100 г/сут и более.
3. Преобладание катаболизма белков над анаболизмом ведет к накоплению
продуктов азотистого об-
мена, в первую очередь мочевины и ее повышен-
ному выведению. Избыток
аминокислот уходит в глю-
конеогенез.
4. Глюкоза и мочевина
осмотически удерживают воду в просвете поччного
канальца и возникает по-
лиурия. Объем мочи воз-растает в 2-3 раза.
5. Повышенный распад
ТАГ в жировой ткани и пе-чени обуславливает ано-
мально высокое окисление
жирных кислот и накопле-ние их недоокисленных
продуктов – кетоновых тел.
Это приводит к кетонемии,
кетонурии и кетоацидозу.
При диабете концентрация кетоновых тел возрастает
в 100-200 раз и достигает
350 мг% (норма 2 мг% или
0,1-0,6 ммоль/л).
6. При полиурии с мо-
чой, кроме воды, теряются соли, в частности карбо-
наты,имеющие щелочнойхарактер. Это усугубляет ацидоз.
7. В результате
.п.п.4,5,6 возникает дегид-
ратация (в тяжелых случаях до5л)организма,которая заключается в падении объ-
ема крови, обезвоживанию клеток и их сморщиванию (дряблая кожа, запавшие гла-
за, мягкие глазные яблоки, сухость слизистых), уменьшению артериального давле-ния. Ацидоз вызывает одышку (дыхание Kussmaul, быстрое и глубокое) и дополни- тельную дегидратацию.
8. Активируется центр жажды и начинается полидипсия.
9. Дегидратация неминуемо приводит к недостаточности кровообращения в тка-
нях – активируется анаэробный гликолиз, накапливается лактат и в дополнение к ке-
тоацидозу возникает лактацидоз.
10. Закисление среды вызывает изменение взаимодействия инсулина с рецепто-
рами, клетки становятся нечувствительными к инсулину – инсулинорезистент-
ность.
11. Ацидоз крови уменьшает концентрацию 2,3-дифосфоглицерата в эритроци-
тах. Это, повышая сродство гемоглобина к кислороду, создает тканевую гипоксию и
усугубляет лактацидоз
Отдаленные последствия
Гипергликемия резко повышает потребление глюкозы инсулиннезависимыми тканями (клетки артериальных стенок, эндотелия, клетки Шванна, эритроциты, хру- сталик и сетчатка глаза, семенники и гломерулярные клетки почек), в них вынужден-но активируется особые пути метаболизма глюкозы. Их интенсивность определяется только доступностью глюкозы:
1. Превращение глюкозы в
сорбитол.
Сорбитол плохо проникает через клеточные мембраны, его накопление в цитозоле приводит
к осмотическому набуханию кле-ток и нарушению их функций. Например, возникновение ката-ракты хрусталика и нейропатий (нарушение осязания) в клетках Шванна
2. Неферментативное гли-козилирование различных бел- ков, изменение их свойств и активация их синтеза за счет избытка энергии:
• увеличивается синтез гликопротеинов базальной мембраны почечных клу-бочков. Это приводит к окклюзии капилляров и нарушению фильтрации • увеличивается синтез гликопротеинов в сетчатке глаза, что вызывает отек
сетчатки и кровоизлияния
• увеличивается синтез гликопротеинов в стекловидном теле
• увеличивается синтез тканевых белков за счет доступности глюкозы и энер-
гии
• гликозилированные белки хрусталика объединяются в крупные агрегаты,
рассеивающие свет. Это вызывает помутнение хрусталика и катаракту.
• гликозилирование гемоглобина в эритроцитах, образование HbA1C • белков свертывающей системы, что увеличивает вязкость крови
• белков ЛПНП, что уменьшает их связывание с рецепторами и повышает концентрацию ХС в крови
• белков ЛПВП, что усиливает их сродство к рецепторам и быструю элимина-
цию из кровотока
Из-за двух последних нарушений возникают макроангиопатии развивается ате-
росклероз сосудов мозга, сердца, почек, конечностей. Характерно в основном для
ИНЗСД.
ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ
При калорийности рациона 2000–3000 ккал суточное потребление углеводов составляет 300-450г. С пищей поступает крахмал, сахароза, лактоза, пищевые волокна (клетчатка и др.). Переваривание углеводов начинается в ротовой полости при участии α-амилазы слюны, которая расщепляет в крахмале α-1,4-гликозидные связи. Полное расщепление крахмала здесь не происходит, так как пребывание пищи во рту кратковременно. Из крахмала в ротовой полости образуются крупные фрагменты – декстрины. Желудочный сок не содержит ферментов, расщепляющих углеводы. Дальнейшее переваривание углеводов происходит в тонком кишечнике. Фермент поджелудочной железы α-амилаза расщепляет α-1,4-гликозидные связи крахмала и декстринов, α-1,6-гликозидные связи расщепляются ферментом кишечного сока – амило-1,6-гликозидазой. При действии двух ферментов образуется дисахарид мальтоза. Амилаза поджелудочной железы не расщепляет β-1.4-гликозидные связи, которыми соединены остатки глюкозы в молекуле целлюлозы. Поэтому пищевые волокна не перевариваются, но они должны присутствовать в рационе, так как улучшают перистальтику, ускоряют чувство насыщения и снижают уровень холестерина в крови, поскольку на них происходит адсорбция желчных кислот и выведение их из организма. Пищевые волокна обязательно должны присутствовать в рационе при ожирении, запорах, атеросклерозе, сахарном диабете. Мальтоза, образовавшаяся из крахмала, а также дисахариды пищи – сахароза и лактоза перевариваются ферментами тонкого кишечника – дисахаридазами. Эти ферменты работают не в просвете кишечника, а на поверхности эпителиальных клеток кишечника. Мальтоза расщепляется мальтазой до 2 молекул глюкозы, лактоза – лактазой до глюкозы и галактозы, сахароза – сахаразой до глюкозы и фруктозы (рис.3). Все моносахариды всасываются, сначала путем облегченной диффузии, а затем активным транспортом в симпорте с ионами Nа+.
Рис.3. Катаболизм дисахаридов и патогенез дисахаридозов
В крови воротной вены содержатся три моносахарида: глюкоза, фруктоза и галактоза. Все они попадают в печень, где происходит унификация фруктозы и галактозы, т.е. они превращаются в глюкозу – единственный моносахарид, используемый всеми клетками нашего организма.
Дисахаридозы – нарушение переваривания дисахаридов, связанные с недостаточной активностью дисахаридаз. Недостаточная активность ферментов может быть врожденной и приобретенной. Симптомы врожденных форм проявляются достаточно рано, например, после первого кормления грудным молоком (при дефиците лактазы) или при добавлении в рацион сахара или крахмала. Приобретенные формы могут наблюдаться при заболеваниях кишечника. Нерасщепленные дисахариды вызывают осмотическую диарею, сбраживаются микрофлорой кишечника с образованием углекислого газа, что приводит к метеоризму, коликам. ОБМЕН ГЛИКОГЕНА
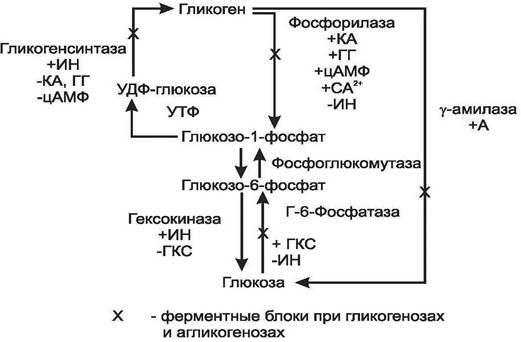
Многие ткани в качестве резервной формы глюкозы синтезируют гликоген. Синтез и распад гликогена обеспечивают постоянство концентрации глюкозы в крови. Синтезгликогена происходит в покое и сытости,как любой анаболический процесс требуетэнергии. Депонируется гликоген главным образом в печени и мышцах. Глюкоза, поступившая в клетку, фосфорилируется при участии гексокиназы за счет АТФ, при этом образуется глюкозо-6-фосфат, который в ходе обратимой реакции под действием фосфоглюкомутазы превращается в глюкозо-1-фосфат. Затем при участии УТФ глюкозо-1-фосфат превращается в УДФ-глюкозу. Эта молекула используется как донор остатков глюкозы при синтезе гликогена.
Так как гликоген в клетке никогда не расщепляется полностью, синтез гликогена осуществляется путем удлинения уже имеющейся молекулы полисахарида, называемой «затравка». К «затравке» последовательно присоединяются остатки глюкозы из УДФ-глюкозы α- 1,4-гликозидной связью при участии фермента гликогенсинтазы. Разветвленная структура гликогена образуется при участии «фермента ветвления» (рис.4). Регуляторными ферментами в синтезе гликогена являются гликогенсинтаза и гексокиназа. Синтез гликогена увеличивается под влиянием инсулина, а тормозится глюкагоном, катехоламинами, глюкокортикостероидами.
Рис.4. Обмен гликогена печени
Распад гликогена происходит путем последовательного отщепления остатков глюкозы в виде глюкозо-1-фосфата. Гликозидная связь расщепляется с присоединением неорганического фосфата, поэтому процесс называется фосфоролизом, а фермент – фосфорилазой. Образовавшийся глюкозо-1-фосфат затем изомеризуется фосфоглюкомутазой до глюкозо-6-фосфата. В печени (но не в мышцах) глюкозо-6-фосфат может гидролизоваться с образованием глюкозы, которая выделяется в кровь. Эту реакцию катализирует глюкозо-6-фосфатаза. Мышечный гликоген не используется для поддержания уровня глюкозы в крови, так как в мышцах нет фермента глюкозо-6-фосфатазы и образование свободной глюкозы там невозможно, а глюкозо-6-фосфат не может проникать через мембрану клеток. Таким образом, печень запасает глюкозу в виде гликогена не столько для собственных нужд, сколько для поддержания постоянной концентрации глюкозы в крови. Функция мышечного гликогена заключается в освобождении глюкозо-6-фосфата, потребляемого в самой мышце для окисления и использования энергии.
Регуляторными ферментами распада гликогена являются фосфорилаза и глюкозо-6-фосфатаза. Процесс распада усиливают катехоламины, глюкагон, глюкокортикостероиды; тормозит инсулин.
|
||
|
|
Последнее изменение этой страницы: 2021-05-12; просмотров: 142; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.41 (0.008 с.) |