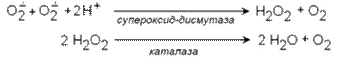Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Транспорт газов эритроцитами.Содержание книги
Поиск на нашем сайте
Основным белком эритроцитов является гемоглобин. Основными гемоглобинами (Hb) взрослого человека являются: гемоглобин А1 (96–98 %) и гемоглобин А2 (1,5-3,5 %). На долю фетального гемоглобина (HbF – α2γ2) приходится 0,5–1 %. Смена гемоглобина F на гемоглобин А происходит во время рождения ребенка, а к 4–6 месяцам жизни уровень фетального гемоглобина составляет 1 %. Благодаря гемоглобину 1л. крови может связывать около 210 мл. кислорода. Связывание кислорода гемоглобином происходит за счёт кооперативного функционирования его субъединиц. Гемоглобин тетрамерный белок и состоит из 2а-субъединиц и 2b-субъединиц, каждая из которых содержит небелковую часть – гем. Гемоглобин осуществляет транспорт кислорода из легких к тканям. При присоединении молекулы кислорода к одной из субъединиц этого тетрамера происходит изменение третичной структуры данной субъединицы. В свою очередь это вызывает целый ряд структурных перестроек в соседних субъединицах, которые приводят к изменению четвертичной структуры. Присоединения последующих молекул кислорода к остальным субъединицам происходит намного эффективнее. В результате присоединения кислорода к гемоглобину эритроцитов в легочных капилярах образуется оксигемоглобин. Отдавая молекулы кислорода оксигемоглобин превращается в дезоксигемоглобин. Высвобождение кислорода тканям зависит от аллостерического регулятора 2,3-дифосфоглицерата в аллостерическом центре молекулы дезоксигемоглобина, при этом сродство гемоглобина к кислороду резко снижается. При переходе в оксигемоглобин 2,3-дифосфоглицерат покидает аллостерический центр. Транспорт СО2 . У взрослого человека при распаде мономеров в тканях образуется до 500л/сут СО2 , который поступает в кровь. При прохождении крови по капиллярам легких происходит диффузия растворенного СО2 из крови через стенки капилляров в просвет легочных альвеол. Только 5-10% СО2 переносится в физически растворенном виде. Попадая в цитоплазму эритроцита СО2 может присоединяться к аминогруппам гемоглобина с образованием карбгемоглобина или вступать в реакцию с водой. Эту реакцию ускоряет Zn2+ - содержащий фермент карбоангидраза. Образовавшаяся угольная кислота диссоциирует на протон и бикарбонат-ион. Избыток НСО3- диффундирует по НСО3/CL- - каналу эритроцита в плазму крови. Протоны, в свою очередь, связываются с оксигемоглобином и облегчают диссоциацию оксигемоглобина с высвобождением кислорода. Перенося кислород, эритроциты его сами не потребляют. Однако незначительная его часть идет на оксисление Fe2+ гемоглобина до Fe3+. В этом случае гемоглобин превращается в метгемоглобин и не способен связывать кислород. В норме в эритроцитах содержится от 0,5 до 3% метгемоглобин. Под воздействием окислителей (метиленового синего и др.) количество метгемоглобина значительно увеличивается (до 30%). Превращение метгемоглобина в гемоглобин происходит с участием фермента метгемоглобинредуктаза и НАДН(НАДФН). При взаимодействии оксида углерода с гемоглобином образуется химическое соединение- карбосигемоглбин, которое неспособно связывать кислород.
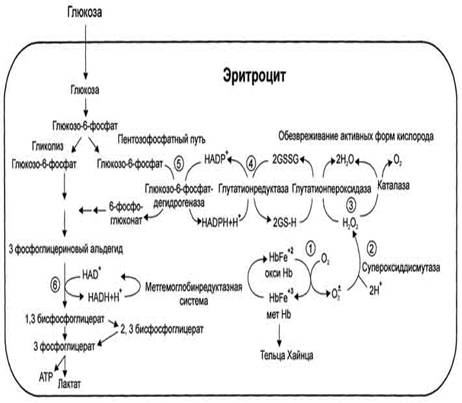
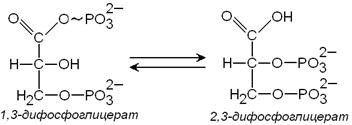
Метаболизм эритроцитов. Обмен глюкозы. Обменные процессы из-за отсутствия органелл в зрелых эритроцитах очень ограничены. 90% глюкозы в эритроцитах распадается в процессе анаэробного гликолиза и 10% окисляется в реакциях пентозофосфатного пути. Конечным продуктом анаэробного гликолиза является молочная кислота, которая выходит в плазму крови и используется другими клетками. Молекулы АТФ, которые образуются в результате анаэробного гликолиза необходимы для активного транспорта ионов через мембрану эритроцита, т.е. для поддержания внутриклеточного градиента концентрации ионов и для поддержания самого гликолиза, требующего затраты АТФ в гексокиназной и фосфофруктокиназной реакциях. Важная особенность анаэробного гликолиза в эритроцитах по сравнению с другими клетками — присутствие в них фермента бисфосфоглицератмутазы, которая катализирует образование 2,3-бисфосфоглицерата из 1,3-бисфосфоглицерата:
| 2,3-бисфосфоглицерат, образуется только в эритроцитах и служит важным аллостерическим регулятором связывания O2 с гемоглобином. В ходе пентозофосфатного цикла вырабатываются восстановленный никотинамидадениндинуклеотид (НАДН+Н+), который используется для восстановления метгемоглобина в гемоглобин, и восстановленный никотинамидадениндинуклеотидфосфат (НАДФН+Н+)- для восстановления глутатиона. Глутатион – это биологически активный трипептид (γ-глутамил-цистеинил-глицил), способный легко окисляться, защищает от окисления и инактивации ряд важных ферментов, содержащих серу, в частности, ферменты, связанные с молекулой гемоглобина и клеточной мембраной.
COOH – CH – CH2 – CH2 – CO – NH – CH – CH2 – SH | | NH2 CO | NH | CH2 Глутатион | COOH Вследствие снижения скорости синтеза АТФ падает активность Na+,K+- АТФ-азы, повышается осмотическое давление и возникает осмотический шок. Дефицит НАДН+Н+ приводит к накоплению метгемоглобина и увеличению образования активных форм кислорода, вызывающих окисление SН-групп в молекулах гемоглобина. Большое содержание кислорода в эритроцитах определяет высокую скорость образования супероксидного анион-радикала O2-, пероксида водорода Н2О2 и гидроксил-радикала ОН'. Постоянным источником активных форм кислорода в эритроцитах является неферментативное окисление железа гемоглобина:
Активные формы кислорода могут вызвать гемолиз эритроцитов. Эритроциты содержат ферментативную систему, предотвращающую токсическое действие радикалов кислорода и разрушение мембран эритроцитов. Восстановленная форма глутатиона (сокращённое обозначение Г-SH) участвует в реакциях обезвреживания пероксида водорода и органических пероксидов (R-O-OH). При этом образуются вода и окисленный глутатион (сокращённое обозначение Г-S-S-Г).
Превращение окисленного глутатиона в восстановленный катализирует фермент глутатионредуктаза.
Антиоксидантные системы имеют для эритроцитов особое значение, так как в эритроцитах не происходит обновления белков путём синтеза. Нарушение любого звена ферментативной системы обезвреживания активных форм кислорода приводит к снижению скорости этого процесса. При генетическом дефекте глюкозо-6-фосфатдегидрогеназы и приеме некоторых лекарств, являющихся сильными окислителями, потенциал глутатионовой защиты может оказаться недостаточным. В норме фермент супероксиддисмутаза катализирует образование пероксида водорода, который под действием глутатионпероксидазы превращается в Н2О. При недостаточной активности ферментов обезвреживания активных форм кислорода происходит окисление SH-групп в остатках цистеина протомеров метгемоглобина и образование дисульфидных связей. Такие структуры называются тельцами Хайнца. Они способствуют разрушению эритроцитов при попадании их в мелкие капилляры. Активные формы кислорода, вызывая перекисное окисление липидов мембран, разрушают мембраны.
Рис.2. Образование и обезвреживание активных форм кислорода в эритроците: 1 - спонтанное окисление Fe2+ в теме гемоглобина - источник супероксидного аниона в эритроцитах; 2 - Супероксиддисмутаза превращает супероксидный анион в пероксид водорода и воду: О2-+ О2- + 2Н+ → Н2О2 + О2; 3 - пероксид водорода расщепляется каталазой: 2Н2О2 → 2Н2О + О2 или глутатионпероксидазой: 2GSH + Н2О2 → GSSG +2Н2О; 4 - Глутатионредуктаза восстанавливает окисленный глутатион: GSSG + NADPH+ + Н+ → 2GSH + NADP+; 5 - NADPH, необходимый для восстановления глутатиона, образуется на окислительном этапе пентозофосфатного пути превращения глюкозы; 6 - NADH, необходимый для восстановления гемоглобина метгемоглобинредуктазной системой, образуется в глицеральдегидфосфатдегидрогеназной реакции гликолиза. Гемоглобинопатии. Известно много генетических вариантов гемоглобина, образование которых приводит к аномалиям эритроцитов или их функции. Среди них наиболее известен гемоглобин S, обусловливающий серповидноклеточную анемию. Серповидноклеточная анемия – наследственное заболевание, обусловленное точечной мутацией гена, кодирующего структуру β – цепи гемоглобина. При этой патологии эритроциты в условиях низкого парциального давления кислорода принимают форму серпа. Гемоглобин S, после отдачи кислорода в тканях превращается в плохо растворимую форму и начинает выпадать в осадок в виде веретенообразных кристаллоидов. Они деформируют клетку и приводят к гемолизу. Болезнь протекает остро, и дети погибают в раннем возрасте. Дефект при серповидноклеточной анемии сводится к замене единственной аминокислоты – глутаминовой в 6 – ом положении с N – конца на валин в β – цепях молекулы гемоглобина. Все остальные аминокислоты распологаются в той же последовательности и в таком же количестве, как и в нормальном гемоглобине Hb A: Hb A: вал – гис – лей - тре – про – глу – глу – лиз…… Hb S: вал – гис – лей - тре – про – вал – глу – лиз…… При этом заболевании отмечают анемию, прогрессирующую слабость, отставание в развитии и желтуху. Талассемии – наследственные заболевания, обусловленное снижением скорости синтеза α– или β– цепей гемоглобина. При β – талассемии не синтезируются β– цепи гемоглобина. Это вызывает образование нестабильных тетрамеров, содержащих только α – цепи. При этом заболевании в костном мозге усиливается разрушение эритробластов, а ускорение разрушения эритроцитов приводит к внутрисосудистому гемолизу. Клинически β – талассемия не проявляется до рождения. В случае α – талассемии нарушается синтез α – цепей гемоглобина F у плода. Наследственный сфероцитоз. Причиной заболевания является дефект белков цитоскелета эритроцитов – спектрина или анкирина, которые обеспечивают поддержание двояковогнутой формы эритроцитов и эластичности мембран. Эритроциты приобретают шарообразную форму, что приводит снижению скорости газообмена. Заболевание сопровождается анемией и желтухой.
2. Лейкоциты (белые кровяные тельца) отличаются большим разнообразием, как по строению, так и по функциональному значению. Лейкоциты являются полноценными клетками с большим ядром, митохондриями и высоким содержанием нуклеиновых кислот. Нормальное количество лейкоцитов в периферической крови – 1,9 – 5,7 х109/л. Регуляция пролиферации стволовой клетки костного мозга в процессе дифференцировки лейкоцитов осуществляется гранулоцитарно-макрофагальными колониестимулирующими факторами. Ингибирование процесса дифференцировки осуществляется клетками, несущими рецепторы к Fc-фрагменту IgG и др. Основной путь получения энергии – аэробный гликолиз. АТФ образуется также в реакциях β-окисления жирных кислот. В лейкоцитах сосредоточен весь гликоген крови, который является источником энергии при недостаточном её поступлении. В лизосомах лейкоцитов локализована мощная система протеолитических ферментов – протеазы, фосфатазы, эстеразы, ДНК-азы, РНК-азы, что обеспечивает участие этих клеток в защитных реакциях организма. Поглощение бактерий лейкоцитами в процессе фагоцитоза сопровождаются резким увеличением потребления кислорода с образованием супероксидного аниона и пероксида водорода, которые оказывают бактерицидное действие. Это явление называется «распираторным взрывом». Лейкоциты активно участвуют в борьбе с чужеродными микроорганизмами. Они устремляются к месту повреждения тканей и накапливаются вблизи него, причем эти клетки мигрирует из кровотока через стенки близлежащих кровеносных сосудов. К месту повреждения их привлекают химические вещества, высвобождаемые поврежденными тканями. Эти клетки способны поглощать бактерии и разрушать их своими ферментами. Таким образом, лейкоциты препятствуют распространению инфекции в организме. Лейкоциты принимают также участие в удалении мертвых или поврежденных тканей. 3.Тромбоциты (кровяные пластинки) - образуются из цитоплазмы мегакариоцитов костного мозга. Это безъядерные фрагменты их цитоплазмы. У здорового человека количество тромбоцитов в периферической крови подвержено значительным колебаниям в течение суток—от 150 до 350 • 109/л. В тромбоцитах протекают основные биохимические процессы: синтез белка, реакции обмена углеводов и липидов, окислительное фосфорилирование
|
||||
|
|
Последнее изменение этой страницы: 2021-01-14; просмотров: 270; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.217.116 (0.012 с.) |

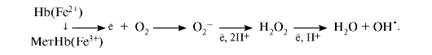

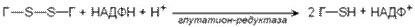 В эритроцитах имеются также ферменты супероксиддисмутаза и каталаза, осуществляющие следующие превращения:
В эритроцитах имеются также ферменты супероксиддисмутаза и каталаза, осуществляющие следующие превращения: