Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Современные представления о сенсорной болевой (ноцицептивной) системеСодержание книги
Поиск на нашем сайте
Болевые рецепторы Болевые раздражения могут возникать в коже, глубоких тканях и внутренних органах. Эти раздражения воспринимаются рецепторами (ноцицепторами), расположенными по всему телу, за исключением головного мозга. Техника микронейрографии сделала возможным утверждать наличие у человека двух таких же типов рецепторов боли (ноцицепторов), как и у других млекопитающих. Анатомически первый тип ноцицепторов представлен свободными нервными окончаниями, разветвлёнными в виде дерева (миелиновые волокна). Они представляют собой быстрые А - дельта волокна, проводящие раздражение со скоростью 6 - 30 м\с. Эти волокна возбуждаются высокоинтенсивными механическими (булавочный укол) и, иногда, термическими раздражениями кожи. А - дельта ноцицепторы располагаются, преимущественно, в коже. Находятся они также и в суставах. Другой тип ноцицепторов представлен плотными некапсулиро-ванными гломерулярными тельцами (немиелиновые С - волокна, проводящие раздражение со скоростью 0,5 - 2 м\с). Эти афферентные волокна у человека и других приматов представлены полимодальными ноцицепторами, поэтому реагируют как на механические, так на температурные и химические раздражения. Они активируются химическими веществами, возникающими при повреждении тканей, являясь одновременно и хеморецепторами. С - волокна распределяются по всем тканям за исключением центральной нервной системы. Однако, они присутствуют в периферических нервах, как nervi nervorum. Волокна, имеющие рецепторы, воспринимающие повреждения тканей, содержат субстанцию Р, выступающую в качестве трансмиттера. Все типы ноцицепторов характеризуются низкой возбудимостью. Биологически активные вещества, взаимодействующие с хемоноцицепторами, называ-ются алгогенами. Выделяют три группы алгогенов: тканевые, плазменные, выделяющиеся из нервных окончаний. К тканевым алгогенам относят серотонин, гистамин, некоторые простогландины, большие концентрации ионов К+, Н+. К плазменным алгогенам относят брадикинин, каллидин, контактный фактор Хагемана. К третьей группе относят олигопептид - субстанцию Р. Задние рога спинного мозга Большинство “болевых волокон” при помощи афферентных нейронов спинномозговых нервов связаны со спинным мозгом или входят в продолговатый мозг в составе тройничного нерва. При вхождении ноцицептивных волокон в спинной мозг, они разделяются на восходящие и нисходящие ветви. Перед своим окончанием в сером веществе задних рогов эти волокна могут направляться к нескольким сегментам спинного мозга. Разветвляясь, они формируют связи с другими многочисленными нервными клетками.. Ноцицептивной информацией прямо или косвенно активируются два основных класса релейных клеток задних рогов спинного мозга: “ноцицептивные специфические” нейроны, активируемые только ноцицептивными стимулами и “wide dynamic range” или “конвергентные” нейроны, активируемые также и не ноцицептивными стимулами. Периферический и центральный контроль локализуется в желатинозной субстанции, примыкающей к клеточному слою.
Восходящие пути боли. Давно известно, что восходящие “болевые пути” находятся в составе переднебоковых канатиков белого вещества спинного мозга и идут контрлатерально стороне вхождения болевых стимулов. Так же хорошо известно, что часть волокон спиноталамического и спиноретикулярного трактов, проводящих болевое раздражение, присутствует в заднебоковом канатике. Спиноталамический тракт может быть, разделён на две части: 1. Неоспиноталамический тракт (быстрое проведение, моносинаптическая передача, хорошо локализованная (эпикритическая) боль, А - волокна). Этот тракт направляется к специфическим латеральным ядрам таламуса (вентро-задне-латеральное и вентрозаднемедиальное ядра). 2. Палеоспиноталамическая система (полисинаптическая передача, медленное проведение, плохо локализованная (протопатическая) боль, С - волокна). Данные пути восходят к неспецифическим медиальным таламическим ядрам (медиальное ядро, интра-ламинарное ядро, срединный центр). На своём пути к медиальным ядрам таламуса тракт направляет часть волокон к ретикулярной формации. Импульсы входящие через неоспиноталамическую систему переключаются на волокна, передающие сигналы к первой соматосенсорной зоне коры, постцентральной извилине и второй соматосенсорной зоне (operculum parietal). Высокая степень топической организации внутри латерального ядра таламуса делает возможным пространственную локализацию боли.
Импульсы, входящие через палеоспиноталамический тракт, переключаются на медиальное ядро таламуса и проецируются на неокортекс диффузным способом. Проекция в лобной области отражает аффективные компоненты боли. Таким образом, в головном мозге нет “болевого центра”, а восприятие и реакция на боль являются функцией ЦНС в целом. Представление об антиноцицептивной системе: характеристика морфофункциональной организации, взаимоотношения с ноцицептивной системой. Опиоидные рецепторы и механизмы Открытие опиоидных пептидов и опиоидных рецепторов относится к началу 70х годов. Клиническое значение имеют три класса опиоидных рецепторов: мю -, каппа - и дельта - рецепторы. Их распределение внутри ЦНС очень вариабильно. Плотное размещение рецепторов обнаружено в задних рогах спинного мозга, в среднем мозге и таламусе. Иммуноцитохимические исследования показали наибольшую концентрацию спинальных опиоидных рецепторов в поверхностных слоях задних рогов спинного мозга. Эндогенные опиоидные пептиды (энкефалин, эндорфин, динорфин) взаимодействуют с опиоидными рецепторами всякий раз, когда в результате преодоления болевого порога возникают болевые раздражения. Факт расположения множества опиоидных рецепторов в поверхностных слоях спинного мозга означает, что опиаты могут легко проникать в него из окружающей спинномозговой жидкости. постцентральная извилина островнковая кора (pars opercularis)
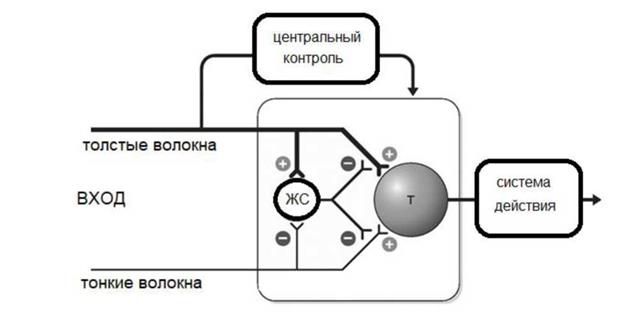
Нисходящая модуляция боли. Известно, что микроинъекции морфина в периакведуктальное серое вещество (PAG) среднего мозга (центральное серое вещество - ЦСВ), также как и его электрическая стимуляция вызывает настолько глубокую анальгезию, что у крыс даже хирургические вмешательства не вызывают каких - либо заметных реакций. Когда были открыты области сосредоточения опиоидных рецепторов и естественных опиатов, стало понятно, что эти отделы ствола мозга являются релейной станцией супраспинальных нисходящих модуляторных контрольных систем. Таким образом, стало ясным, что эндогенные опиоиды играют важную роль в болевом контроле как на спинальном, так и супраспинальном уровне. На спинальном уровне энкефалины представляют собой основные нейромедиаторы, ингибирующие афферентные волокна, где медиатором является субстанция Р. На супраспинальном уровне эндорфины и энкефалины вовлечены в нисходящих путях. Кроме опиатного контроля болевой чувствительности известен серотонинэргический и адренэргический механизмы контроля. Свидетельства существования нисходящего серотонинергического контроля на ноцицептивные спинальные нейроны широко представлены в результатах исследования на животных. Описанные выше механизмы модуляции боли объединены в так называемую антиноцицептивную систему. Функция этой системы состоит в том, чтобы контролировать ноцицептивную систему, обеспечивая оптимальный уровень ее активации. Современные теории боли В 1965 г. R.Melzack, P.Wall предложили теорию входных ворот, которая, в настоящее время принята большинством исследователей. (Рис.2.)
Рис.2. Теория воротного контроля. ЖС - желатинозная субстанция, Т - нейроны первого центрального переключения.
Первое положение теории входных ворот заключается в том, что передача нервных импульсов от афферентных волокон к нейронам спинного мозга, передающим сигналы в головной мозг, модулируется спинальным воротным механизмом - системой интернейронов, локализованных в желатинозной субстанции (вторая и третья пластины заднего рога) спинного мозга. Установлено, что нейроны этих пластин получают аксонные терминали от множества афферентных волокон большого и малого диаметра, в эту же область входят дендриты более глубоких пластин задних рогов спинного мозга. Это дало основание полагать, что желатинозная субстанция действует как спинальные ворота, модулируя проведение нервных импульсов от периферических рецепторов к нейронам спинного мозга, дающим проекции в головной мозг, - релейным нейронам. Согласно второму положению теории входных ворот, спинальный воротный механизм регулируется относительно большим количеством импульсов, поступающих по афферентным волокнам большого и малого диаметра. Воротный механизм ограничивает передачу нервных импульсов к релейным нейронам при высокой интенсивности импульсации по афферентным волокнам большого диаметра (закрывает ворота) и, наоборот, облегчает прохождение нервных импульсов к релейным нейронам в случаях, когда возрастает афферентный поток по афферентным волокнам малого диаметра (открывает ворота). Третье положение теории R.Melzack и P.Wall заключается в том, что спинальный воротный механизм находится под влиянием нервных импульсов, которые передаются по волокнам нисходящих систем из коры большого мозга и ствола мозга. Такие когнитивные (оценивающие) факторы, как внимание, тревога и др., оказывают мощное влияние на процесс восприятия боли. Четвертое положение теории состоит в том, что специализированная система волокон большого диаметра, обладающая высокой скоростью проведения, является триггером (пусковым устройством) центрального контроля, который активирует избирательные когнитивные процессы влияющие через нисходящие волокна на модулирующие свойства спинального воротного механизма. Функция триггера центрального контроля, по мнению R.Melzack, P.Wall несут система задние столбы - медиальная петля и дорсолатеральная система. Пятое положение теории следующее: когда возбуждение релейных нейронов спинного мозга превышает критический уровень, то их импульсация приводит к возбуждению ноцицептивной системы. При достижении критического уровня возбуждения в релейных нейронах поток импульсов передается в мозг, главным образом через антеролатеральную восходящую систему: волокнами неоспино-таламического пути в
вентробазальный таламус и далее в соматосенсорную кору большого мозга, а также волокнами палеоспиноталамического пути в ретикулярную формацию, медиальный интраламинарный таламус и структуры лимбической системы. Активация этих структур мозга является основой сенсорных, мотивационных и когнитивных компонентов общей реакции на боль. Таким образом, основные принципы теории входных ворот учитывают специфичность рецепторов, физиологические механизмы конвергенции, суммации, облегчения и торможения, роль восходящих и нисходящих систем головного и спинного мозга. Лекция 26 ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ЦЕЛЕНАПРАВЛЕННОГО План лекции 1. Физиология высшей нервной деятельности - научное направление Российской физиологической школы в изучении целенаправленного поведения животных и человека. 2. Понятие условного и безусловного рефлексов. Классификация условных рефлексов. Условия выработки условных рефлексов. 3. Системность в работе головного мозга. Представление о динамическом стереотипе. 4. Теории формирования временных связей, лежащих в основе выработки условных рефлексов. 5. Корковое торможение: условия возникновения, классификация, вероятные механизмы. Физиология высшей нервной деятельности - научное направление Одним из механизмов приспособления животных и человека к изменяющимся условиям существования во внешней среде является деятельность нервной системы, в основе которой лежат рефлекторные механизмы. В процессе эволюции возникли наследственно закрепленные реакции (безусловные рефлексы), которые объединяют и согласовывают функции различных органов, осуществляют адаптацию организма. У человека и высших животных в процессе индивидуальной жизни возникают качественно новые рефлекторные реакции, которые И. П. Павлов назвал условными рефлексами, считая их самой совершенной формой приспособления. Именно условные рефлексы, как полагал И.П.Павлов являются функциональной единицей высшей нервной деятельности (ВНД). Высшая нервная деятельность - это совокупность безусловных и условных рефлексов, а также высших психических функций, которые обеспечивают адекватное поведение в изменяющихся природных и социальных условиях. Впервые предположение о рефлекторном характере деятельности высших отделов мозга было высказано И.М.Сеченовым, что позволило распространить рефлекторный принцип и на психическую деятельность человека. Идеи И.М.Сеченова получили экспериментальное подтверждение в трудах И.П.Павлова, который разработал метод объективной оценки функций высших отделов мозга - метод условных рефлексов. Понятие условного и безусловного рефлексов. Классификация
И.П.Павлов показал, что все рефлекторные реакции можно разделить на две группы: безусловные и условные. Сравнительная характеристика безусловных и условных рефлексов представлена в таблице 1 Таблица 1 СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА БЕЗУСЛОВНЫХ И
Безусловные рефлексы могут быть простыми и сложными. Сложные врожденные безусловно-рефлекторные реакции называются инстинктами. Их характерной особенностью является цепной характер реакций. Условный рефлекс - это сложная многокомпонентная реакция, которая вырабатывается на базе безусловных рефлексов с использованием предшествующего индифферентного раздражителя. Он имеет сигнальный характер, и организм встречает воздействие безусловного раздражителя подготовленным. Например, у спортсмена в предстартовый период происходит перераспределение крови, усиление дыхания и кровообращения, и когда мышечная нагрузка начинается, организм уже к ней подготовлен. Правила выработки условных рефлексов Для выработки условного рефлекса необходимо: 1. Наличие двух раздражителей, один из которых безусловный (пища, болевой раздражитель и др.), вызывающий безусловно-рефлекторную реакцию, а другой - условный (сигнальный), сигнализирующий о предстоящем безусловном раздражении (свет, звук, вид пищи и т.д.); 2. Многократное сочетание условного и безусловного раздражителей (хотя, при определенных условиях, иногда возможно образование условного рефлекса при их однократном сочетании); 3. Условный раздражитель должен предшествовать действию безусловного; 4. В качестве условного раздражителя может быть использован любой раздражитель внешней или внутренней среды, который должен быть по возможности индифферентным, не вызывать обронительной реакции, не обладать чрезмерной силой. Вместе с тем индифферентный раздражитель должен восприниматься организма, т.е. он должен быть пороговым; 5. Безусловный раздражитель должен быть достаточно сильным, значимым для организма, в противном случае временная связь не сформируется; 6. Необходимо устранить посторонние раздражители, так как они могут вызывать внешнее торможение условного рефлекса; 7. Животное, у которого вырабатывается условный рефлекс, должно быть здоровым; 8. При выработке условного рефлекса должна быть выражена мотивация, например, при выработке пищевого слюноотделительного рефлекса животное должно быть голодным, у сытого - этот рефлекс не вырабатывается. Классификация условных рефлексов 1. Условные рефлексы, также как и безусловные можно классифицировать по биологической модальности - пищевые, питьевые, оборонительные; 2. В зависимости от характера взаимосвязи сигнального, условного и безусловного раздражителей, условные рефлексы делятся на натуральные и искусственные. Натуральные условные рефлексы вырабатываются на агенты, которые в естественных условиях являются свойством безусловного раздражителя, действуют вместе с раздражителем, вызывающим безусловный рефлекс (например, вид пищи, ее запах и т.д.). Все остальные условные рефлексы искусственные, т.е. вырабатываются на агенты, в норме не связанные с действием безусловного раздражителя, например, пищевой слюно-отделительный рефлекс на звонок. 3. По эффекторному признаку условные рефлексы делят на секреторные, двигательные, сердечные, сосудистые и т.д. 4. По роли в реализации целенаправленного поведения условные рефлексы делят на подготовительные и исполнительные. 5. Если выработать прочный условный пищевой рефлекс, например, на свет, то такой рефлекс является условным рефлексом первого порядка. На его базе можно выработать условный рефлекс второго порядка, для этого дополнительно применяют новый, предшествующий сигнал, например звук, подкрепляя его условным раздражителем первого порядка (светом). В результате нескольких сочетаний звука и света звуковой раздражитель также начинает вызывать слюноотделение. Таким образом возникает новая более сложная опосредованная временная связь. Следует подчеркнуть, что подкреплением для условного рефлекса второго порядка является именно условный раздражитель первого порядка, а не безусловный раздражитель (пища), так как если и свет и звук подкреплять пищей, то возникнут два отдельных условных рефлекса первого порядка. При достаточно прочном условном рефлексе второго порядка можно выработать условный рефлекс третьего порядка. Для этого используется новый раздражитель, например, прикосновение к коже. В этом случае прикосновение подкрепляется только условным раздражителем второго порядка (звуком), звук возбуждает зрительный центр, а последний - пищевой центр. Возникает еще более сложная временная связь. Рефлексы более высокого порядка (4, 5, 6 и т.д.) вырабатываются только у приматов и человека. 6. По характеру отношения животного или человека к безусловному раздражителю, на базе которого вырабатывают условный рефлекс, условные рефлексы делят на положительные и отрицательные. Положительные условные рефлексы сближают с безусловным раздражителем. Отрицательные уловные рефлексы либо удаляют от него, либо препятствуют сближению. 7. В зависимости от длительности периода изолированного действия условного сигнала (ПИД) условные рефлексы делят на совпадающие (ПИД = от 0,5 до 3,0 сек.), короткоотставленные (ПИД = от 3,0 до 30 сек.), нормальноотставленные (ПИД = от 30 до 60 сек.), запаздывающие (ПИД = более 60 сек.). Период изолированного действия - отрезок времени от начала действия условного сигнала до момента действия безусловного раздражителя. Системность в работе головного мозга. Представление о Отдельные условные рефлексы в определенной ситуации могут связываться между собой в комплексы. Если осуществлять ряд условных рефлексов в строго определенном порядке, с примерно одинаковыми временными интервалами, и весь этот комплекс сочетаний многократно повторять, то в мозге сформируется единая система, имеющая специфическую последовательность рефлекторных реакций, т.е. ранее разрозненные рефлексы связываются в единый комплекс. Нейроны головного мозга, обладая большой функциональной подвижностью, тем не менее, могут стойко удерживать систему ответных реакций на повторяющиеся условные раздражения. Возникает динамический стереотип, который выражается в том, что на систему различных условных сигналов, действующих всегда один за другим через определенное время, вырабатывается постоянная и прочная система ответных реакций. В дальнейшем, если применять только первый раздражитель, то в ответ будут развиваться все остальные реакции. Динамический стереотип - характерная особенность психической деятельности человека. Многие наши навыки, например, способность писать, играть на музыкальных инструментах, танцевать и т.д., являются автоматическими цепями двигательных актов. В процессе жизни человека обычно вырабатываются и более сложные стереотипы поступков: поведение после пробуждения или перед сном, режим труда, отдыха, питания. Возникают относительно устойчивые формы поведения в обществе, во взаимоотношениях с другими людьми, в оценке текущих событий и реагирования на них. Такие стереотипы имеют большое значение в жизни человека, так как позволяют выполнять многие виды деятельности с меньшим напряжением нервной системы. Биологический смысл динамических стереотипов сводится к тому, чтобы освободить корковые центры от решения стандартных задач, для того чтобы обеспечить выполнение более сложных, требующих эвристического мышления. Вместе с тем, если стереотипы поведения формировались многие годы, их переделка в изменяющихся условиях окружающей среды затруднена. Ломка старых стереотипов является стрессогенной ситуацией и может привести к невротическим состояниям. Теории формирования временных связей, лежащих в основе 1. В какой части головного мозга происходит образование временной связи? Физиологической основой для возникновения условных рефлексов служит образование функциональных временных связей в высших отделах ЦНС. Временная связь - это совокупность нейрофизиологических, биохимических и ультраструктурных изменений в мозге, возникающих в процессе совместного действия условного и безусловного раздражителей. И.П.Павлов высказал предположение, что при выработке условного рефлекса происходит формирование временной нервной связи между двумя группами клеток коры - корковыми представительствами условного и безусловного рефлексов. Возбуждение от центра условного рефлекса может передаваться к центру безусловного рефлекса от нейрона к нейрону. Следовательно, первый путь образования временной связи между корковыми представительствами условного и безусловного рефлексов является внутрикортикальным. Однако при разрушении коркового представительства условного рефлекса выработанный условный рефлекс сохраняется. По-видимому, образование временной связи идет не только между корковыми представительствами условного и безусловного сигналов, но и между подкорковым центром условного рефлекса и корковым центром безусловного рефлекса. 2. Какие процессы в головном мозгу обеспечивают формирование временных связей? Имеются различные мнения по вопросу о механизмах образования временной связи. А) Биохимическая теория объясняет механизм формирования временных связей за счет синтеза белковых веществ в нейронах, которые обеспечивают условный рефлекс. При введении животным в желудочки мозга веществ, блокирующих синтез белка, выработка условных рефлексов нарушается. Поскольку синтез белка обеспечивается системой ДНК - РНК, следовательно в процессе выработки условных рефлексов принимают участие механизмы генетической памяти, локализованные в ядерном аппарате нейронов. Современные исследования показали, что белковые вещества, синтезируемые в ходе выработки условного рефлекса, обеспечивают сближение возбужденных нейронов и активацию их синаптических механизмов. Б) Анатомическая теория объясняет механизмы формирования временной связи структурной перестройкой синаптического аппарата нервных клеток, участвующих в формировании условных рефлексов. Электрономикроскопические исследования показали, что при выработке условных рефлексов у экспериментальных животных в нейронах ассоциативной коры происходит активация ряда ранее неактивных синапсов. В) Теория конвергентного замыкания временных связей на отдельных нейронах П.К.Анохина. В соответствии с представлениями П.К.Анохина формирование временных связей происходит в мультиконвергентных нейронах. Мультконвергентные нейроны - это такие нейроны, на тело которых сходятся различные по модальности сенсорные потоки. Например, афферентные возбуждения от рецепторов зрительной сенсорной системы и от вкусовых рецепторов. Мультиконвергентные нейроны расположены в ассоциативной коре, в стволовой части мозга. Процессы, протекающие в мультконвергентных нейронах описаны биохимической и анатомической теориями.
|
||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2020-12-17; просмотров: 197; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.147.78.185 (0.016 с.) |