Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Проводящие пути головного и спинного мозгаСодержание книги
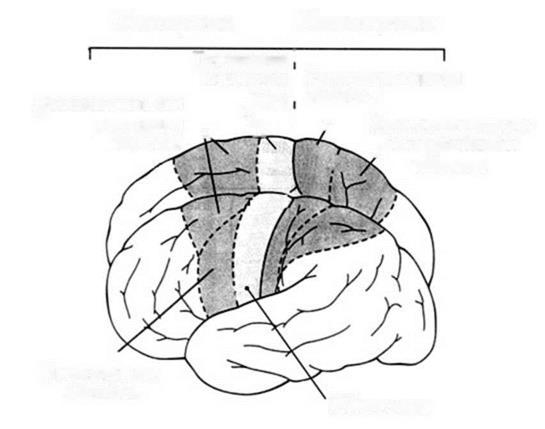
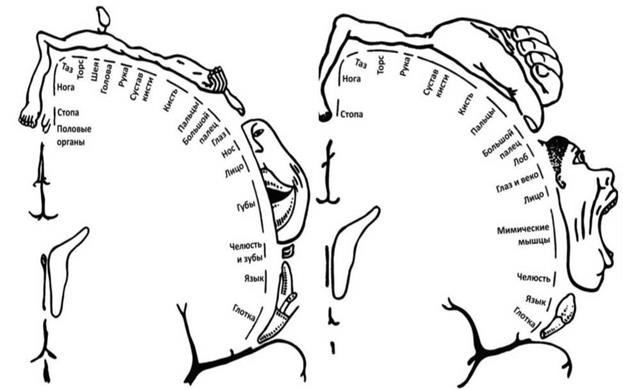
Поиск на нашем сайте Системы нервных волокон, проводящих импульсы от кожи и слизистых оболочек, внутренних органов и органов движения к различным отделам спинного и головного мозга, в частности к коре полушарий большого мозга, называются восходящими, или чувствительными, афферентными, проводящими путями. Системы нервных волокон, передающих импульсы от коры или нижележащих ядер головного мозга через спинной мозг к рабочему органу (мышце, железе и др.), называются двигательными, или нисходящими, эфферентными, проводящими путями. Проводящие пути образованы цепями нейронов, причем чувствительные пути обычно состоят из трех нейронов, а двигательные - из двух. Первый нейрон всех чувствительных путей располагается всегда вне мозга, находясь в спинномозговых узлах или чувствительных узлах черепных нервов. Последний нейрон двигательных путей всегда представлен клетками передних рогов серого вещества спинного мозга или клетками двигательных ядер черепных нервов. Чувствительные пути. Спинной мозг проводит четыре вида чувствительности: тактильную (чувство прикосновения и давления), температурную, болевую и проприоцептивную (от рецепторов мышц и сухожилий, так называемое суставно-мышечное чувство, чувство положения и движения тела и конечностей). Основная масса восходящих путей проводит проприоцептивную чувствительность. Это говорит о важности контроля движений, так называемой обратной связи, для двигательной функции организма. Путь болевой и температурной чувствительности - латеральный спиноталамический путь. Первым нейроном этого пути являются клетки спинномозговых узлов. Аксоны данных нейронов образуют задние корешки и идут в спинной мозг, оканчиваясь на клетках задних рогов (2-й нейрон). Отростки вторых нейронов через комиссуру спинного мозга переходят на противоположную сторону (образуют перекрест) и поднимаются в составе бокового канатика спинного мозга в продолговатый мозг. Там они примыкают к медиальной чувствительной петле и идут через продолговатый мозг, мост и ножки мозга к латеральному ядру таламуса, где переключаются на 3-й нейрон. Отростки клеток ядер таламуса образуют таламокортикальный пучок, проходящий через заднюю ножку внутренней капсулы к коре постцентральной извилины (область чувствительного анализатора). В результате того что волокна по пути перекрещиваются, импульсы от левой половины туловища и конечностей передаются в правое полушарие, а от правой половины - в левое. Передний спиноталамический путь состоит из волокон, проводящих тактильную чувствительность, он проходит в переднем канатике спинного мозга. Пути мышечно-суставной (проприоцептивной) чувствительности направляются к коре полушарий большого мозга и в мозжечок, который участвует в координации движений. К мозжечку идут два спиномозжечковых пути - передний и задний. Задний спиномозжечковый путь (Флексига) начинается от клетки спинномозгового узла (1-й нейрон). Периферический отросток входит в состав спинномозгового нерва и заканчивается рецептором в мышце, капсуле суставов или связках. Центральный отросток в составе заднего корешка входит в спинной мозг и заканчивается в клетках ядра, расположенного у основания заднего рога (2-й нейрон). Отростки вторых нейронов поднимаются в дорсальной части бокового канатика этой же стороны и через нижние ножки мозжечка идут к клеткам коры червя мозжечка. Волокна переднего спиномозжечкового пути (Говерса) образуют перекрест дважды; в спинном мозге и в области верхнего паруса, а затем через верхние ножки мозжечка достигают клеток коры червя мозжечка. Проприоцептивный путь к коре больших полушарий представлен двумя пучками: нежным (тонким) и клиновидным. Нежный пучок (Голля) проводит импульсы от проприорецепторов нижних конечностей и нижней половины тела и лежит медиально в заднем канатике. Клиновидный пучок (Бурдаха) примыкает к нему снаружи и несет импульсы от верхней половины туловища и от верхних конечностей. Второй нейрон этого пути лежит в одноименных ядрах продолговатого мозга. Их отростки образуют перекрест в продолговатом мозге и соединяются в пучок, называемый медиальной чувствительной петлей. Она доходит до латерального ядра таламуса (3-й нейрон). Отростки третьих нейронов через внутреннюю капсулу направляются в чувствительную и частично двигательную зоны коры. Двигательные пути представлены двумя группами. 1. Пирамидные (кортико-спинальный и кортико-ядерный, или кортикобульбарный) пути, проводящие импульсы от коры к двигательным клеткам спинного и продолговатого мозга, являющиеся путями произвольных движений. 2. Экстрапирамидные, рефлекторные двигательные пути, входящие в состав экстрапирамидной системы. Пирамидный, или кортико-спинальный путь начинается от больших пирамидных клеток (Беца) коры верхних 2/3 предцентральной извилины и околоцентральной дольки, проходит через внутреннюю капсулу основание ножек мозга, основание моста, пирамиды продолговатого мозга. На границе со спинным мозгом он разделяется на боковой и передний пирамидные пучки. Боковой (большой) образует перекрест и спускается в боковом канатике спинного мозга, заканчиваясь на клетках переднего рога. Передний не перекрещивается и идет в переднем канатике. Образуя посегментный перекрест, его волокна также заканчиваются на клетках переднего рога. Отростки клеток переднею рога образуют передний корешок, двигательную порцию спинномозгового нерва и заканчиваются в мышце двигательным окончанием. Кортико-ядерный путь начинается в нижней трети предцентральной извилины, идет через колено (изгиб) внутренней капсулы и заканчивается на клетках двигательных ядер черепных нервов противоположной стороны. К рефлекторным двигательным путям (экстрапирамидным) относятся красноядерно-спинномозговой (руброспинальный) путь - от клеток красного ядра среднего мозга, тектоспиналъный путь - от ядер холмиков пластинки крыши среднего мозга (четверохолмия), связанный со слуховыми и зрительными восприятиями, и вестибуло-спинальный - от вестибулярных ядер из ромбовидной ямки, связанный с поддержанием равновесия тела. 2. Физиология конечного мозга Конечный мозг, или полушария большого мозга, достигшие своего наивысшего развития у человека, справедливо считается самым сложным и самым удивительным созданием природы. Функции этого отдела центральной нервной системы настолько отличаются от функций ствола и спинного мозга, что они выделяются в особую главу физиологии, называемую высшей нервной деятельностью. Этот термин введен И. П. Павловым. Деятельность нервной системы, направленную на объединение и регуляцию всех органов и систем организма, И. П. Павлов назвал низшей нервной деятельностью. Под высшей нервной деятельностью он понимал поведение, деятельность, направленную на приспособление организма к изменяющимся условиям внешней среды, на уравновешивание с окружающей средой. Большие достижения И. П. Павлова в области изучения функций полушарий большого мозга объясняются тем, что он доказал рефлекторную природу деятельности коры и открыл присущий только ей новый, тип рефлексов, а именно условных рефлексов. Открыв основной механизм деятельности коры полушарий большого мозга, он тем самым создал объективный метод изучения ее функций - метод условных рефлексов. Как выяснилось в дальнейшем, условные рефлексы есть те элементарные акты, те "кирпичики", из которых строится поведение животных и человека. Значение полушарий у различных животных до И. П. Павлова изучали путем хирургического удаления их. Результаты удаления полушарий большого мозга птиц и собак показали, что вегетативные функции: кровообращение, дыхание, пищеварение и др., существенно не нарушаются. При тщательном уходе животное живет долго. Но при этом, нарушается его связь с внешней средой. На непосредственно действующие раздражители - укол булавкой, раздражение слизистой оболочки рта пищей - возникает вполне адекватная реакция: лапа отдергивается, пища проглатывается, т. е. у животного сохраняются врожденные безусловные рефлексы. Но утрачиваются все приобретенные реакции поведения, все выработанные в процессе индивидуальной жизни условные рефлексы. Для изучения локализации (места нахождения) функций в коре полушарий большого мозга, или, иными словами, значения отдельных зон коры, применяют различные методы: частичное удаление коры, электрическое и химическое раздражение, запись биоэлектрической активности мозга, метод условных рефлексов и другие. Метод раздражения позволил установить в коре следующие зоны: двигательные (моторные), чувствительные (сенсорные) и ассоциативные. Двигательные зоны коры Движения возникают при раздражении коры в области предцентральной извилины. Электрическое раздражение верхней части извилин вызывает движение мышц ног и туловища, средней - рук, нижней - мышц лица. Величина корковой двигательной зоны пропорциональна не массе мышц, а точности движений. Особенно велика зона, управляющая движениями кисти руки, языком, мимической мускулатурой лица (рис. 5). В V слое коры двигательных зон обнаружены гигантские пирамидные клетки (пирамиды Беца), отростки которых спускаются к двигательным нейронам среднего, продолговатого и спинного мозга, иннервирующим скелетную мускулатуру. Путь от коры к двигательным нейронам носит название пирамидного пути. Это путь регуляции произвольных движений. После повреждения моторной зоны произвольные движения не могут осуществляться. Раздражение моторной зоны сопровождается движениями на противоположной половине тела, что объясняется перекрестом пирамидных путей на их пути к двигательным нейронам, иннервирующим мышцы. Сенсорные зоны коры Экстирпация (удаление) различных участков коры у животных позволила в общих чертах установить локализации сенсорных (чувствительных) функций. Затылочные доли оказались связанными со зрением, височные - со слухом. Зона коры, куда проецируется данный вид чувствительности, называется первичной проекционной зоной. Кожная чувствительность человека, чувства прикосновения, давления, холода и тепла проецируются в постцентральную извилину. В верхней ее части находится проекция кожной чувствительности ног и туловища, ниже - рук и совсем внизу - головы (рис. 5). Абсолютная величина проекционных зон отдельных участков кожи неодинакова. Так, например, проекция кожи кисти рук занимает в коре большую площадь, чем проекция поверхности туловища (рис. 5). Величина корковой проекции пропорциональна значению данной рецептивной поверхности в поведении. Суставно-мышечная, проприоцептивная, чувствительность проецируется в постцентральную и предцентральную извилины.
Рис 5. Схема двигательного (справа) и чувствительного (слева) гомункулусов Показаны проекции частей тела человека на область коркового конца двигательного и соматосенсорного анализаторов. (Проекции моторного и чувствительного гомункулусов представлены и в левом и в правом полушариях). Зрительная зона коры находится в затылочной доле. При раздражении ее возникают зрительные ощущения - вспышки света; удаление ее приводит к слепоте. Удаление зрительной зоны на одной половине мозга вызывает слепоту на одной половине каждого глаза, так как каждый зрительный нерв делится в области основания мозга на две половины (образует неполный перекрест), одна из них идет к своей половине мозга, а другая - к противоположной. При повреждении наружной поверхности затылочной доли не проекционной, а ассоциативной зрительной зоны зрение сохраняется, но наступает расстройство узнавания (зрительная агнозия). Больной, будучи грамотным, не может прочесть написанное, узнает знакомого человека после того, как тот заговорит. Способность видеть - это врожденное свойство, но способность узнавать предметы вырабатывается в течение жизни. Бывают случаи, когда от рождения слепому возвращают зрение уже в старшем возрасте. Он еще долгое время продолжает ориентироваться в окружающем мире на ощупь. Проходит немало времени, пока он научится узнавать предметы с помощью зрения. Функция слуха обеспечивается височными долями больших полушарий. Раздражение их вызывает простые слуховые ощущения. Удаление обеих слуховых зон вызывает глухоту, а одностороннее удаление понижает остроту слуха. При повреждении участков коры слуховой зоны может наступить слуховая агнозия: человек слышит, но перестает понимать значение слов. Родной язык становится ему так же непонятен, как и чужой, иностранный, ему незнакомый. Заболевание носит название слуховой агнозии. Обонятельная зона коры находится на основании мозга, в области парагиппокампальной извилины. Проекция вкусового анализатора, по-видимому, находится в нижней части постцентральной извилины, куда проецируется чувствительность полости рта и языка. Применение современных микроэлектродных методов для изучения функций корковых нейронов в значительной мере расширило представления о переработке сенсорной информации в неокортексе. В 1957 г. американский исследователь В. Маунткасл, анализируя ответы клеток в соматосенсорной (сенсомоторной) коре кошки на стимулы различных модальностей, обнаружил следующий интересный факт. При погружении микроэлектрода перпендикулярно поверхности соматосенсорной коры все встречаемые им клетки отвечали на раздражитель одной и той же модальности, например на легкое прикосновение к коже или на движение в суставе. Если же электрод погружали под углом к поверхности коры, то на его пути попадались нейроны с различной сенсорной модальностью, чередовавшиеся с определенной периодичностью. На основании этих экспериментальных фактов был сделан вывод о том, что соматосенсорная кора организована в элементарные функциональные единицы — колонки, ориентированные перпендикулярно поверхности. Диаметр такой колонки определяется горизонтальным распространением терминалей афферентного таламокортикального волокна и вертикальной ориентацией дендритов пирамидных клеток. Колонка является элементарным блоком сенсомоторной коры, где осуществляется локальная переработка информации от рецепторов одной модальности. Эта гипотеза колончатой организации неокортекса получила широкое распространение и дала толчок к дальнейшим исследованиям в этой области. Согласно современным представлениям, каждая функциональная колонка сенсомоторной коры состоит из нескольких морфологических микромодулей, объединяющих пять—шесть гнездообразно расположенных нейронов. В его состав входит несколько пирамидных клеток, апикальные дендриты которых максимально сближены и образуют дендритный пучок; в пределах этого пучка возможны электротонические связи, которые обеспечивают, по всей вероятности, синхронную работу всего объединения. 3. Физиология лимбической системы В конечном мозге располагаются образования (поясная извилина, гиппокамп, миндалевидное тело, область перегородки), составляющие лимбическую систему. Они участвуют в поддержании постоянства внутренней среды организма, регуляции вегетативных функций и формировании эмоций и мотиваций. Эту систему иначе называют "висцеральным мозгом", так как эта часть конечного мозга может рассматриваться как корковое представительство интерорецепторов. Сюда поступает информация от внутренних органов. При раздражении желудка, мочевого пузыря в лимбической коре возникают вызванные потенциалы. Электрическое раздражение различных областей лимбической системы вызывает изменения вегетативных функций: кровяного давления, дыхания, движений пищеварительного тракта, тонуса матки и мочевого пузыря. Разрушение отдельных частей лимбической системы приводит к нарушению поведения: животные могут становиться более спокойными или, напротив, агрессивными, легко дающими реакции ярости, изменяется половое поведение. Лимбическая система имеет широкие связи со всеми областями головного мозга, ретикулярной формацией и гипоталамусом. Она обеспечивает высший корковый контроль всех вегетативных функций (сердечно-сосудистой, дыхательной, пищеварительной, обмена веществ и энергии).
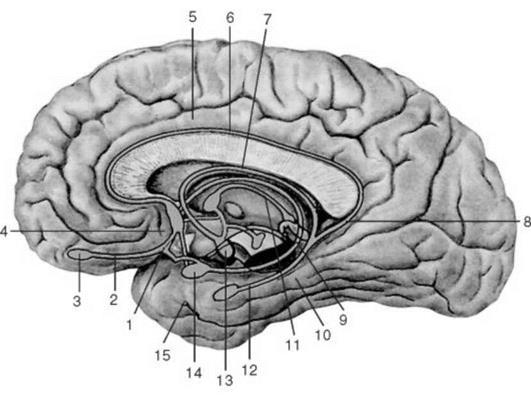
Рис 6. Образования головного мозга, относящиеся к лимбической системе. 1 - обонятельный треугольник; 2 - обонятельный тракт; 3 - обонятельная луковица; 4 - паратерминальная извилина; 5 - поясная извилина; 6 - серый покров; 7 - свод; 8 - перешеек поясной извилины; 9 - терминальная пластинка; 10 - парагиппокампальная извилина; 11 - мозговая полоска таламуса; 12 - гиппокамп; 13 - сосцевидное тело; 14 - миндалевидное тело; 15 - крючок Ассоциативные зоны коры Проекционные зоны коры занимают в мозге человека небольшую долю всей поверхности коры. Остальная поверхность занята так называемыми ассоциативными зонами. Нейроны этих областей не связаны ни с органами чувств, ни с мышцами, они осуществляют связь между различными областями коры, интегрируя объединяя все притекающие в кору импульсы в целостные акты научения (чтение, речь, письмо), логического мышления, памяти и обеспечивая возможность целесообразной реакции поведения. При нарушениях ассоциативных зон появляются агнозии - неспособность узнавания и апраксии - неспособность производить заученные движения. Например, стереоагнозия выражается в том, что человек не может найти на ощупь у себя в кармане ни ключа, ни коробки спичек, хотя зрительно он их сейчас же узнает. При нарушении ассоциативных зон коры может наступить афазия - потеря речи. Афазия может быть моторной и сенсорной. Моторная афазия возникает при поражении задней трети нижней лобной извилины слева, так называемого центра Брока (этот центр находится только в левом полушарии). Больной понимает речь, но сам говорить не может. При сенсорной афазии, поражении центра Вернике в задней части верхней височной извилины, больной речи не понимает. При аграфии человек разучивается писать, при апраксии - производить заученные движения: зажечь спичку, застегнуть пуговицу, пропеть мелодию и др. Изучение локализации функции методом условных рефлексов на живом здоровом животном позволило И. П. Павлову обнаружить факты, на основе которых им была построена теория динамической локализации функций в коре, затем блестяще подтвержденная при помощи микроэлектродного исследования нейронов. У собак вырабатывали условные рефлексы, например на зрительные раздражения - свет, различные фигуры - круг, треугольник, а затем удаляли всю затылочную, зрительную, зону коры. После этого условные рефлексы исчезали, но проходило время, и нарушенная функция частично восстанавливалась. Это явление компенсации, или восстановления, функции И. П. Павлов объяснил, высказав мысль о существовании ядра анализатора, расположенного в определенной зоне коры, и рассеянных клеток, разбросанных по всей коре, в зонах других анализаторов. За счет этих сохранившихся рассеянных элементов происходит восстановление утраченной функции. Собака может отличать свет от тьмы, но тонкий анализ, установление различий между кругом и треугольником, ей недоступен, он свойствен только ядру анализатора. Микроэлектродное отведение потенциалов от отдельных нейронов коры подтвердило наличие рассеянных элементов. Так, в двигательной зоне коры обнаружили клетки, дающие разряд импульсов на зрительные, слуховые, кожные раздражения, а в зрительной зоне коры выявлены нейроны, отвечающие электрическими разрядами на осязательные, звуковые, вестибулярные и обонятельные раздражители. Кроме того, были найдены нейроны, которые отвечают не только на "свой" раздражитель, как теперь говорят, раздражитель своей модальности, своего качества, но и на один - два чужих. Их назвали полисенсорными нейронами. Динамическая локализация, т. е. способность одних зон замещаться другими, обеспечивает коре больших полушарий высокую надежность. Лекция 13 ГУМОРАЛЬНЫЕ ВЗАИМОСВЯЗИ В ОРГАНИЗМЕ План лекции 1. Представления о гуморальных механизмах регуляции. 2. Понятие гормона, классификация гормонов, механизмы действия на клетки мишени. 3. Понятие о железах внутренней секреции, классификация желез внутренней секреции. 4. Регуляция активности желез внутренней секреции. В процессе эволюции при образовании многоклеточных организмов возникла необходимость возникновения механизмов, обеспечивающих взаимосвязь, как между отдельными клетками тканей, так и отдельными органами в целом организме. В результате эволюционного отбора сформировались два механизма объединения (интеграции) элементов в целое - механизмов нервной и гуморальной регуляции. О механизмах нервной регуляции мы говорили с вами на предыдущих лекциях. Сегодня поговорим о механизмах гуморальной регуляции. Гуморальная регуляция - совокупность физиологических, биохимических и биофизических механизмов, обеспечивающих изменение состояния отдельных клеток, тканей, органов и систем посредством химических соединений через внутреннюю среду организма. Понятие внутренней среды организма ввел известный французский физиолог Клод Бернар. Под внутренней средой организма он понимал совокупность биологических жидкостей - кровь, лимфу, тканевую жидкость. Химические вещества, которые могут включаться в механизмы гуморальной регуляции можно разделить на три класса: 1) Растворимые в воде соли или электролиты; 2) Продукты метаболизма или метаболиты. 3) Биологически активные вещества, включая гормоны; К первой группе можно отнести соединения калия, натрия, кальция, магния. Впрочем, в организме человека можно обнаружить почти все известные на сегодняшний день элементы. Поэтому каждое из этих соединений при увеличении или уменьшении его содержания во внутренней среде организма в той или иной степени влиять на состояние клеток, тканей, органов или систем. Ко второй группе химических соединений относят диоксид углерода (СО2), угольную кислоту (Н2СО3), молочную кислоту, пировиноградную кислоту, продукты расщепления АТФ (АДФ, неорганический фосфор и др.). К третьей группе относят гормоны и парагормоны (тканевые гормоны). Термин гормон был предложен английскими физиологами В.Бейлисом и Е.Старлингом в 1905 году. Термин гормон происходит от греческого термина hormao - привожу в движение, побуждаю. Гормонами называют высокоактивные химические вещества, образующиеся и выделяющиеся специализированными клетками, тканями или органами во внутреннюю среду организма. Гормоны отличаются от других веществ, оказывающих свое влияние гуморально, по следующим критериям: 1) Гормоны образуются специализированными клетками; 2) Гормоны оказывают свое влияние дистантно, по принципу «Всем! Всем! Всем!», но взаимодействуя лишь с клетками-мишенями. 3) Гормоны обладают чрезвычайно высокой биологической активностью. Например, вам известно, что адреналин увеличивает частоту и силу сердечных сокращений. Согласно расчетам, 1 грамм адреналина способен активировать 100 000 000 изолированных сердец лягушек. Вам известен гормон инсулин, который участвует в регуляции обмена глюкозы. 1 грамм инсулина способен понизить уровень глюкозы в крови у 125 000 кроликов. Источники гормонов Источниками гормонов могут являться: 1) Эндокринные специализированные железы, например щитовидная железа, надпочечники, околощитовидная железа; 2) Эндокринная ткань в органе. Например, эндокринная ткань, содержащая а и в клетки, в поджелудочной железе. 3) Рассеянные клетки, обладающие эндокринной функцией. Например, G клетки пилорического отдела желудка, вырабатывающие гормон гастрин. 4) Клетки, которые наряду с эндокринной функцией выполняют еще и другие специализированные функции. Например, нервные клетки гипоталамуса (промежуточный мозг), выполняют свои функции нервных клеток (генерация потенциала действия и проведение его к другим нервным клеткам). Кроме того, нервные клетки этой области способные синтезировать и секретировать гормоны. Например, антидиуретический гормон или вазопрессин и окцитоцин. Первый гормон принимает участие в регуляции объема выделяемой во внешнюю среду мочи, второй - в регуляции сократительной активности гладкой мускулатуры матки. Классификация гормонов 1. Деление гормонов на группы по критерию химического строения. По данному критерию выделяют три группы гормонов.
А) Гормоны, производные аминокислот. Например, адреналин, гормон тирозина. Йод содержащие гормоны щитовидной железы также являются производными от аминокислоты тирозина. Б) Гормоны белковой природы. В данной группе выделяют три подгруппы: - олигопептиды, белки с малой молекулярной массой, например, антидиуретический гормон (синоним вазопрессин), окситоцин, гастрин и другие; - собственно белки, например, инсулин, глюкагон и другие; - глюкопротеиды, например, фоликулостимулирующий гормон, лютеинизирующий гормон. В) Стероидные гормоны. К этой группе относят гормоны коры надпочечников, половые гормоны. 2. Деление гормонов по механизму действия. По данному критерию гормоны делят на две группы: А) Гормоны мембранного типа действия, например, инсулин, глюкагон. Рецепторы, чувствительные к гормонам этого типа встроены в биологическую мембрану клеток мишеней. Поэтому гормон «находит» рецептор на мембране клетки-мишени и взаимодействует с ним. Далее гормон-рецепторный комплекс активирует вторичные посредники, которые сопрягают мембранные процессы с цитоплазматическими. В роли вторичных посредников могут выступать те же вещества, что участвуют при реализации передачи сигнала через синапс: - аденилатциклаза - циклический аденозин-монофосфат (ЦАМФ); - гуанилатциклаза - циклический гуанозин-монофосфат (ЦГМФ); - фосфолипаза С - инозитол-3-фосфат; - Са++ - кальмодулин. Б) Гормоны внутриклеточного (цитозольного) типа действия. К этой группе относят стероидные гормоны, половые гормоны. Рецепторы, чувствительные к гормонам данного типа находятся внутри клеток. Поэтому гормоны этого типа должны проникнуть внутрь клетки, провзаимодействовать с рецептором, и только после этого они могут оказать свой эффект на клетку. Гормоны цитозольного типа действия оказывают свой эффект через наследственный аппарат клетки, находящийся в ядре. 3. Классификация гормонов по физиологическому эффекту. По этому критерию гормоны делят на: - Эффекторные, т.е. оказывающие свой эффект на клетку мишень тканей и органов; - Тропные или гормоны-регуляторы. Представители этой группы синтезируются в передней доле гипофиза. Основное назначение данных гормонов - регуляция активности гипофиз-зависимых желез внутренней секреции. К таким гормонам относят адренокортикотропный гормон, тиреотропный гормон, гонадотропный гормон. - Нейрогормоны (регуляторы-регуляторов). К этой группе относят рилизинг-гормоны (либерины и статины), которые вырабатываются в гипоталамической области. Основное назначение гормонов этой группы - регуляция активности клеток аденогипофизи по выработке тропных гормонов. Синтез гормонов эндокринными клетками происходит непрерывно. Его интенсивность зависит не только от регуляторных сигналов звена управления, но и от величины секреции, т.е. регуляция активности желез внутренней секреции осуществляется как по возмущению, так и по отклонению. Известный в биохимии принцип торможения конечным продуктом обусловливает подавление активности процессов синтеза гормона при его накоплении в продуцирующих клетках. Выделение гормона из клеток желез внутренней секреции приводит к увеличению его синтеза. Активность желез внутренней секреции может также зависеть от концентрации гормона во внутренней среде организма. При высокой концентрации гормона во внутренней среде организма активность продуцирующих клеток снижается. Указанное обстоятель-ство требует осторожного использования гормонов с прикладными целями. Например, в спорте получило широкое применение анаболиков стероидной природы, препаратов, способствующих увеличению мышечной массы. Однако, в силу того, что анаболики этой группы по своей структуре близки к мужским половым гормонам, это может привести у мужчин к гипофункции половых желез с последующей их гипотрофией и даже атрофией. Наконец, в ряде случаев, количество синтезируемого и выделяемого во внутреннюю среду организма гормона зависит от количества во внутренней среде субстрата, содержание которого регулируется данным гормоном. Например, активность Р клеток поджелудочной железы по синтезу и выделению во внутреннюю среду организма инсулина зависит от концентрации глюкозы: повышение содержания глюкозы во внутренней среде приводит к усилению синтеза и секреции р клетками инсулина. Транспорт гормонов. Транспорт гормонов осуществляется, как правило, в связанной форме. Этот процесс осуществляется либо специальными белками переносчиками, которые циркулируют во внутренней среде организма, либо на поверхности мембраны клеток. Например, в роли переносчиков могут выступать эритроциты - клетки крови. Типы действия гормонов на клетки мишени 1) Метаболическое действие. В этом случае гормоны вызывают изменение обмена веществ и энергии в клетках-мишенях. Данные изменения могут быть связаны с увеличением проницаемости биологических мембран, увеличением синтеза каких-либо ферментов или изменения их активности. Например, выраженным метаболическим эффектом обладают гормоны щитовидной железы. 2) Морфогенетическое действие. В этом случае гормоны влияют на процессы изменения клеток мишеней в ходе индивидуального развития человека. Например, соматотропин, гормон передней доли гипофиза оказывает влияние на рост человека. Половые гормоны участвуют в формировании вторичных половых признаков. 3) Кинетическое действие. Это действие гормонов связано с их способностью запускать те или иные физиологические процессы. Например, окситоцин, гормон гипоталамической области активирует сокращение гладкой мускулатуры матки, а антидиуретический гормон включает механизм обратного всасывания воды в почках. 4) Коррегирующее действие. Это действие гормонов связано с их способностью модулировать текущую активность клеток мишеней, органов и тканей. Например, изменение частоты сердечных сокращений под действием адреналина. 5) Реактогенное действие. Это действие гормонов связано с их способностью менять реакцию клеток, тканей и органов на различные раздражители, в т.ч. на действие других гормонов. Например, гастрин и холецистокинин изменяют возбудимость нервных клеток центров голода и насыщения находящихся в гипоталамической области. Понятие о железах внутренней секреции, классификация желез внутренней секреции Железы внутренней секреции - это железы, не имеющие выводных протоков для выделения синтезируемых ими биологически активных веществ. Железы внутренней секреции выделяют синтезируемые ими биологически активные вещества во внутреннюю среду для распространения их по всему организму. 1). Железы внутренней секреции по критерию их локализации делят на центральные и периферические. К центральным относят эпифиз, гипоталамическую область, гипофиз. К периферическим железам внутренней секреции относят щитовидную, паращитовидную железы, надпочечники, островковый аппарат поджелудочной железы, половые железы, вилочковую железу и др. 2). По критерию механизма управления железы внутренней секреции делят на гипофиз зависимые и гипофиз независимые. К гипофиз зависимым железам внутренней секреции относят щитовидную железу, корковый слой надпочечников, половые железы, эпифиз. К гипофиз независимым железам внутренней секреции относят паращитовидную железу, островковый аппарат поджелудочной железы, мозговой слой надпочечников, вилочковую железу. Регуляция активности желез внутренней секреции Регуляция активности желез внутренней секреции осуществляется по отклонению и возмущению. Механизмы регуляции по отклонению мы уже рассматривали на сегодняшней лекции при обсуждении вопроса регуляции активности гормон продуцирующих клеток. Регуляция активности желез внутренней секреции по возмущению осуществляется нервным и гуморальным путем. Например, активация мозгового слоя надпочечников осуществляется при возбуждении симпатического отдела автономной нервной системы, с участием задних ядер гипоталамической области промежуточного мозга. Гуморальный путь активации гипофиз зависимых желез внутренней секреции начинается также в гипоталамической области, где выделяются соответствующие рилизинг-гормоны - либерины. Эти гормоны активируют клетки переднего отдела гипофиза, которые продуцируют тропные гормоны, а те в свою очередь, активируют гипофиз зависимые железы внутренней секреции. Например, такой путь активации можно указать по отношению к корковому слою надпочечников. Воздействие на организм сильного раздражителя приводит в состояние возбуждения не только симпато-адреналового комплекса, но и активирует нейросекреторную активность клеток гипоталамуса и в, частности, способствует выделению гормона кортиколиберина. Кортиколиберин с током крови гипофизарной системы кровообращения попадает в переднюю долю гипофиза, где активирует синтез и выделение адренокортикотропного гормона (АКТГ). Этот гормон с током крови разносится по всему организму и «находит» клетки мишени в корковом слое надпочечников, которые отвечают на данное воздействие активацией синтеза и секреции кортикостероидов, в частности, гидрокортизона. Оба представленных выше механизма регуляции активности желез внутренней секреции взаимодополняют друг друга на разных этапах приспособления организма к изменяющимся условиям окружающей среды. Лекция №14 ФИЗИОЛОГИЯ КРОВИ План лекции 1. Понятие крови. Состав крови. Функции крови. 2. Современные представления об иммунологических свойствах крови, учение о группах крови, правила переливания крови. 3. Гемостаз, виды гемостаза. Понятие крови. Состав крови. Функции крови. Кровь - это составная часть внутренней среды организма. Нередко кровь рассматривают как разновидность ткани, отличительной особенностью которой является преобладание количества межклеточной жидкости над клеточными элементами, в связи с чем, кровь является разновидностью биологической жидкости. Соотношение клеточных элементов к жидкой части называется гематокритом. Клеточных элементов в крови содержится около 40-48%, а жидкой части - 52 - 60% от всего объема крови. Жидкая часть крови получила название плазмы. Клеточный состав крови (форменные элементы крови). Клеточный состав крови представлен эритроцитами, лейкоцитами, и тромбоцитами. Эритроциты - безъядерные клеточные образования, содержащие белок гемоглобин, придающий эритроцитам красную окраску. Этот белок принимает участие в транспорте газов - кислорода и углекислого газа. В крови у человека содержится от 4500000 до 5000000 эритроцитов в 1 мм3 или 4.5 - 5.0 х 1012/л. Лейкоциты - клетки «белого» ростка крови. В крови их содержится от от 6000 до 8000 в 1 мм3 или 6 - 8 х 109/л. Состав лейкоцитов неоднороден - 63% составляют гранулосодержащие лейкоциты (гранулоциты), а 37% составляют клетки, в цитоплазме которых не содержатся гранулы (агранулоциты). Среди грануло
|
|||||||||
|
|
Последнее изменение этой страницы: 2020-12-17; просмотров: 194; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.137 (0.053 с.) |