Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Белки, синтезирующиеся в печени
Дефицит α1-антитрипсина относится к генетическим наследственным заболеваниям. Гаптоглобин представляет собой гликопротеин, состоящий из полипептидных цепей α и β, которые ковалентно соединены между собой дисульфидными связями. Гаптоглобин образуется преимущественно в гепатоцитах. Низкий уровень гаптоглобина наблюдается при тяжёлом хроническом заболевании печени и гемолитическом кризе. Церулоплазмин (ферро-О2-оксидоредуктаза) представляет собой мультифункциональный медьсодержащий белок, осуществляющий неспеци-фическую антиоксидантную защиту организма. При болезни Вильсона уровень церулоплазмина снижен. Низкая концентрация церулоплазмина наблюдается при тяжёлом декомпенсированном циррозе печени другой этиологии (не связанной с болезнью Вильсона). Высокий уровень церулоплазмина можно выявить у беременных, при лечении эстрогенами и при обструкции крупных жёлчных протоков. Уровень компонента С3 комплемента при циррозе печени снижен, при хроническом гепатите в пределах нормы, а при компенсированном билиарном циррозе повышен. Снижение содержания компонента С3 комплемента объясняется также повышенным потреблением белков системы комплемента вследствие её активации. Преходящее снижение уровня компонента С3 комплемента обнаруживают на ранней иммунокомплексной стадии острого гепатита В. Альфа-фетопротеин (АФП) ‒ белок сыворотки крови развивающегося эмбриона человека. У взрослых он почти полностью исчезает из крови вскоре после рождения, но появляется при развитии гепатоцеллюлярной карциномы (ГЦК), а также раке яичка и яичников. Экспрессия гена АФП в печени происходит при процессах некроза и воспаления в печени, сопровождающихся нарушением межклеточного взаимодействия гепатоцитов. В наибольшей степени клеточно-матриксные взаимодействия в печени нарушаются при ГЦК, что подтверждается тем, что самые высокие сывороточные уровни АФП регистрируются именно при этой патологии, а его концентрация зависит от объема и темпов роста опухоли. Кроме этого, повышенные уровни АФП являются фактором риска развития ГЦК у больных циррозом печени. Повышение уровня АФП характерно и для ЦП, поскольку при этом заболевании также нарушаются клеточно-матриксные взаимодействия гепатоцитов вследствие усиления фиброзообразования в печени.
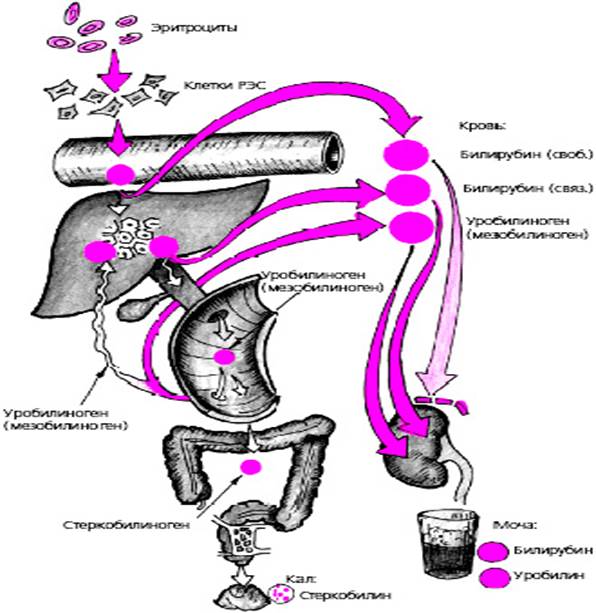
Углеводный обмен. Печень играет основную роль в обмене углеводов. Механизмы его нарушения при циррозе сложны и раскрыты не полностью. У больных циррозом печени при исследовании натощак уменьшается роль углеводов как источника энергии и увеличивается доля жиров. Это может быть обусловлено уменьшением образования глюкозы печенью или снижением запаса гликогена в печёночной ткани. После приёма пищи у больных циррозом печени, как и у здоровых людей, отмечается быстрая утилизация пищевых углеводов, которая выражена даже в большей степени из-за нарушения способности печени к их депонированию. Это сопровождается мобилизацией триглицеридов в качестве источника энергии Пигментный обмен. Обмен билирубина Билирубин - тетрапиррольный пигмент с молекулярной формулой C33H56N406 и молекулярной массой 584,65 Д. Он образуется в процессе катаболизма геминовой части гемоглобина (протопорфирина IX) эритроцитов, закончивших свой жизненный цикл. Кроме того, билирубин может образоваться при катаболизме других гемсодержащих белков (миоглобин, каталаза, пероксидаза), правда, в значительно меньших количествах. При средней продолжительности жизни эритроцитов 120 дней в сутки в организме формируется билирубин в количестве 3,8±0,6 мг/кг массы тела. Эритроцит подвергается разрушению главным образом в клетках РЭС печени и селезенки, лимфатических узлов и костного мозга. При старении эритроцитов снижается содержание сиаловых кислот в составе гликопротеинов цитоплазматической мембраны. Измененные углеводные компоненты гликопротеинов мембран эритроцитов связываются с рецепторами клеток РЭС, и эритроциты поглощаются клетками путем эндоцитоза. Процессу распада подвергается содержащийся в эритроцитах гемоглобин. Вначале происходит разрыв метанового мостика между I и II пиррольными ядрами порфиринового кольца с одновременным окислением Fe2+ в Fe3+ при участии ферментативного комплекса гемоксигеназы. Образовавшийся таким образом пигмент получил название вердоглобин. Дальнейшие превращения приводят к потере вердоглобином железа и глобина. Железо при этом пополняет запасы депо, глобин гидролизуется ферментами лизосом с образованием свободных аминокислот, использующихся организмом для синтеза белка. Порфириновое же кольцо гема разворачивается в цепь с формированием желчного пигмента зеленого цвета - биливердина. Биливердин восстанавливается при участии фермента биливердинредуктазы в основной и важнейший красно-желтый пигмент желчи - билирубин. При распаде 1 г гемоглобина образуется 35 мг билирубина.
Этот образовавшийся в клетках РЭС билирубин: Нерастворим в воде, поэтому не проходит через почечный фильтр; 1. Хорошо растворим в липидах, поэтому: • легко проникает через липидный слой мембран клеток; • способен накапливаться в тканях, богатых липидами (особенно в нервной ткани); 2. Обладает выраженной токсичностью, так как нарушает процессы окислительного фосфорилирования в клетках, нарушает синтез белка, поток ионов калия через мембраны клеток. 3. Непрямой, так как дает реакцию с диазореактивом только после перехода в растворимое соединение при добавлении акселератора (кофеина). Билирубин, образовавшийся вне печени, циркулирует в крови в нековалентной связи с альбумином. Это препятствует обратной диффузии билирубина в ткани и, возможно, способствует его целенаправленному поступлению в печень. Некоторые эндогенные и экзогенные вещества способны вытеснять билирубин из его связи с альбумином. Билирубин, связанный с альбумином, попадает в печени через поры эндотелиальных клеток в пространство Диссе и непосредственно контактирует с синусоидальной мембраной гепатоцитов. В мембрану встроены транспортные белки для билирубина, которые облегчают его поступление в клетку путем диффузии. Транспортная функция самого важного в количественном отношении транспортного белка зависит как от ионов натрия, так и от ионов хлора. Для данного белка характерна кинетика насыщения, и он обеспечивает транспорт как непрямого, так и прямого билирубина. За этот транспортный белок конкурируют лекарственные препараты и другие экзогенные вещества. Билирубин, поступивший внутрь клетки, связывается с белками. Таким образом может обеспечиваться его накопление в нетоксичной форме и предотвращаться его обратная диффузия в кровь. Самым важным внутриклеточным белком связывания является лигандин - изофермент или субъединица глутатиона-S-трансферазы. Конъюгация билирубина в печеночных клетках представляет собой главный этап в обмене билирубина и служит предпосылкой его последующей экскреции с желчью. При конъюгации оба остатка пропионовой кислоты билирубина подвергаются этерификации с глюкуроновой кислотой. При этом вначале возникает моноглюкуронид, а затем - билирубин-диглюкуронид. Неконъюгированный билирубин представляет собой неполярное (жирорастворимое) вещество. В реакции конъюгации он превращается в полярное (водорастворимое вещество) и может благодаря этому выделяться в жёлчь. Эта реакция протекает с помощью микросомального фермента уридиндифосфатглюкуронилтрансферазы (УДФГТ), превращающего неконъюгированный билирубин в конъюгированный моно- и диглюкуронид билирубина. УДФГТ является одной из нескольких изоформ фермента, обеспечивающих конъюгацию эндогенных метаболитов, гормонов и нейротрансмиттеров.
Ген УДФГТ билирубина находится на 2-й паре хромосом. Структура гена сложная. У всех изоформ УДФГТ постоянными компонентами являются экзоны 2 - 5 на 3'-конце ДНК гена. Для экспрессии гена необходимо вовлечение одного из нескольких первых экзонов. Детали строения гена важны для понимания патогенеза неконъюгированной гипербилирубинемии (синдромы Жильбера и Криглера-Найяра), когда в печени содержание ферментов, ответственных за конъюгацию, снижено или они отсутствуют. Активность УДФГТ при печёночно-клеточной желтухе поддерживается на достаточном уровне, а при холестазе даже увеличивается. У новорождённых активность УДФГТ низкая. У человека в жёлчи билирубин представлен в основном диглюкуронидом. Превращение билирубина в моноглюкуронид, а также в диглюкуронид происходит в одной и той же микросомальной системе глюкуронилтрансферазы. При перегрузке билирубином, например при гемолизе, образуется преимущественно моноглюкуронид, а при уменьшении поступления билирубина или при индукции фермента возрастает содержание диглюкуронида. Наиболее важное значение имеет конъюгация с глюкуроновой кислотой, однако небольшое количество билирубина конъюгируется с сульфатами, ксилозой и глюкозой; при холестазе эти процессы усиливаются. В поздних стадиях холестатической или печёночно-клеточной желтухи, несмотря на высокое содержание в плазме, билирубин в моче не выявляется. Очевидно, причиной этого является образование билирубина типа III, моноконъюгированного, который ковалентно связан с альбумином. Он не фильтруется в клубочках и, следовательно, не появляется в моче. Это снижает практическую значимость проб, применяемых для определения содержания билирубина в моче. Экскреция билирубина в канальцы происходит с помощью семейства АТФ-зависимых мультиспецифичных транспортных белков для органических анионов. Скорость транспорта билирубина из плазмы в жёлчь определяется этапом экскреции глюкуронида билирубина. Жёлчные кислоты переносятся в жёлчь с помощью другого транспортного белка. Наличие разных механизмов транспорта билирубина и жёлчных кислот можно проиллюстрировать на примере синдрома Дубина-Джонсона, при котором нарушается экскреция конъюгированного билирубина, но сохраняется нормальная экскреция жёлчных кислот. Большая часть конъюгированного билирубина в жёлчи находится в смешанных мицеллах, содержащих холестерин, фосфолипиды и жёлчные кислоты. Значение аппарата Гольджи и микрофиламентов цитоскелета гепатоцитов для внутриклеточного транспорта конъюгированного билирубина пока не установлено.
Далее конъюгированный билирубин вместе с другими компонентами желчи поступает в тонкий кишечник, поступившие билирубинглюкурониды гидролизуются специфическими бактериальными ферментами (β-глюкуронидазами, которые гидролизуют связь между билирубином и остатком глюкуроновой кислоты). Освободившийся в ходе этой реакции билирубин под действием кишечной микрофлоры восстанавливается с образованием бесцветных тетрапиррольных соединений - уробилиногенов. Втонком кишечнике часть уробилиногена (мезобилиноген -15-20%) вновь всасывается и с кровью воротной вены поступает в печень, где окисляется с образованием пирролов, которые выводятся далее с желчью в кишечник. Поэтому в физиологических условиях уробилиногена нет ни в общей системе кровотока, ни в моче. В толстом кишечнике уробилиноген под действием ферментов микрофлоры кишечника переходит в стеркобилиноген. В нижних отделах толстого кишечника часть стеркобилиногена всасывается стенкой кишечника и по системе геморроидальных вен попадает в общую систему кровотока и оттуда в мочу. Так называемый уробилин мочи, в сущности, представляет собой преимущественно окисленный стеркобилиноген, а при заболеваниях печени к последнему добавляется уробилиноген из-за неспособности печени к его утилизации. На воздухе оба соединения быстро окисляются, переходя в окрашенные в красно-коричневый цвет продукты, обозначаемые общим термином - уробилиновые тела (уробилиноиды). В толстом кишечнике большая часть стеркобилиногена окисляется до стеркобилина, обусловливающего нормальную окраску кала.
Рис.6. Обмен билирубина в норме (источник gepatit5neo.ru)
Дифференциальная диагностика желтух. Возрастание уровня общего билирубина в сыворотке крови до уровня выше 20,5 мкмоль/л называется гипербилирубинемией. Это состояние может быть следствием образования билирубина в большем количестве, чем то, которое нормальная печень может экскретировать; повреждений печени, нарушающих экскрецию билирубина в нормальных количествах, а также вследствие закупорки желчевыводящих протоков печени, что препятствует выведению билирубина. Во всех этих случаях билирубин накапливается в крови и по достижении определенных концентраций диффундирует ткани, окрашивая их в желтый цвет. Это состояние называется желтухой. В зависимости от того, какой тип билирубина присутствует в сыворотке крови -неконъюгированный (непрямой) или конъюгированный (прямой), гипербилирубинемия классифицируется как неконъюгированная I и регургитационная (коньюгированная) соответственно. В клинической практике наиболее широкое распространение получило деление желтух на гемолитические, паренхиматозные и обтурационные. Гемолитические и паренхиматозные желтухи - это неконъюгированная, а обтурационные конъюгированная гипербилирубинемия. В некоторых случаях желтуха может быть смешанной по патогенезу.
Увеличение содержания билирубина в крови может обусловливаться следующими причинами: 1. увеличение интенсивности гемолиза эритроцитов; 2. поражение паренхимы печени с нарушением ее билирубинвыделительной функции; 3. нарушение оттока желчи из желчных путей в кишечник; 4. выявление ферментного звена, обеспечивающего биосинтез глюкуронидов билирубина. 5. нарушение печеночной секреции конъюгированного (прямого) билирубина в желчь. Гемолитическая желтуха. Увеличение интенсивности гемолиза наблюдается при гемолитических анемиях. Гемолиз также может быть усилен при В12-дефицитных анемиях, малярии, массивных кровоизлияниях в ткани, легочных инфарктах, при синдроме размозжения (неконъюгированная гипербилирубинемия). В результате усиленного гемолиза происходит интенсивное образование в ретикулоэндотелиальных слетках свободного билирубина из гемоглобина. В то же время печень оказывается неспособной к образованию столь большого количества билирубинглюкуронидов, что и приводит к накоплению свободного билирубина (непрямого) в крови и тканях. Однако даже при значительном гемолизе неконъюгированная гипербилирубинемия обычно незначительна (менее 68,4 мкмоль/л) вследствие большой способности печени к конъюгированию билирубина. Помимо увеличения уровня общего билирубина, при гемолитической желтухе повышается выделение уробилиногена с мочой и калом, так как он образуется в кишечнике в большом количестве. Наиболее частой формой неконъюгированной гипербилирубинемии является «физиологическая желтуха» у новорожденных. Причинами ее являются гемолиз эритроцитов и незрелое состояние печеночной системы поглощения, конъюгации (сниженная активность УДФ-глюкуронилтрансферазы) и секреции билирубина. В связи с тем, что билирубин, накапливающийся в крови находится в неконъюгированном (свободном) состоянии, когда его концентрация в крови превышает уровень насыщения альбумина (34.2-42,75 мкмоль/л), он способен преодолевать гематоэнцефалический барьер. Это может привести к гипербилирубинемической токсической энцефалопатии. При гепатоцеллюлярной (паренхиматозной) желтухе, наступает деструкция гепатоцитов, нарушается экскреция прямого (конъюгированного) билирубина в желчные капилляры, и он попадает непосредственно в кровь, где содержание его значительно увеличивается. Крометого, снижается способность печеночных клеток синтезировать билирубин-глюкурониды, вследствие чего количество непрямого билирубина также увеличивается. Повышение концентрации в крови прямого билирубина приводит к его появлению в моче вследствие фильтрации через мембрану почечных клубочков. Непрямой билирубин, несмотря на увеличение концентрации в крови в мочу не поступает. Поражение гепатоцитов сопровождается нарушением их способностиразрушать до ди- и трипирролов всосавшийся из тонкого кишечника мезобилиноген (уробилиноген). Повышение содержания уробилиногена в моче может наблюдаться еще в дожелтушный период. В разгар вирусного гепатита возможно снижение и даже исчезновение уробилиногена в моче. Это объясняется тем, что увеличивающийся застой желчи в печеночных клетках ведет к уменьшению выделения билирубина и, следовательно, к уменьшению образования уробилиногена в желчевыводящих путях. В дальнейшем, когда функция печеночных клеток начинает восстанавливаться, желчь выделяется в большом количестве, при этом снова появляется уробилиноген в больших количествах, что в данной ситуации расценивается как благоприятный прогностический признак. Стеркобилиноген попадает в большой круг кровообращения и выделяется почками с мочой в виде уробилина. Основными причинами паренхиматозных желтух являются острые и хронические гепатиты, циррозы печени, токсичные вещества (хлороформ, четыреххлористый углерод, ацетаминофен), массивное распространение в печени раковой опухоли, альвеолярный эхинококк и множественные абсцессы печени. При вирусных гепатитах степень билирубинемии в какой-то мере коррелирует с тяжестью заболевания. Так, при гепатите В при легкой форме течения заболевания содержание билирубина не выше 90 мкмоль/л (5 мг%), при среднетяжелой - в пределах 90-170 мкмоль/л (5-10 мг%), при тяжелой - свыше 170 мкмоль/л (выше 10 мг%). При развитии печеночной комы билирубин может повышаться до 300 мкмоль/л и более. Однако следует иметь в виду, что степень повышения билирубина в крови не всегда зависит от тяжести патологического процесса, а может быть обусловлена темпами развития вирусного гепатита и печеночной недостаточности. Обтурационная (обструктивная, подпеченочная) желтуха обусловлена обструкцией вне- или внутрипеченочных желчных путей, которая вызывает частичное или полное прекращение оттока желчи. При обтурации общего желчного протока (камень, воспаление, опухоль и т. д) из-за скопления Таблица 5.
|
|||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2020-11-11; просмотров: 82; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.86.134 (0.02 с.) |