Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Цитогенетическая характеристика синдромов Прадера-Вилли и АнгельманаСодержание книги
Поиск на нашем сайте
Наиболее убедительные данные получены при синдроме Прадера-Вилли (СПВ) и синдроме Ангельмана (СЭ), которые, имея существенно разные клинические проявления, в своей основе имеют сходные молекулярно-цитогенетические изменения (табл. 2 и 3).
Таблица 2 Корреляция генотип-фенотип при синдроме Ангельмана
Таблица 3Корреляция генотип-фенотип при синдроме Прадера-Вили
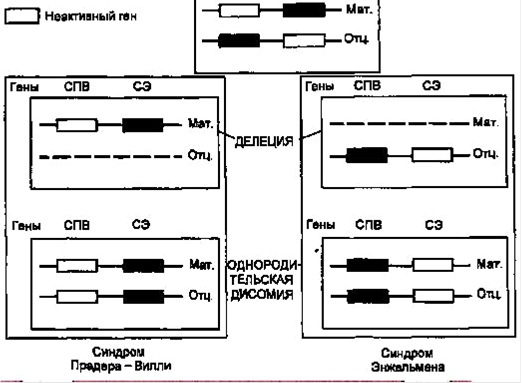
Наиболее частой причиной возникновения СПВ и СА является протяженная (до 4 млн п.н.) деления критического района 15(q11-q13),которую находят у 70-75 % больных с этими синдромами. Делецию при СПВ обнаруживают на отцовской хромосоме 15,а при СЭ делеция той же области на ее материнском гомологе. Второй причиной возникновения СПВ и СЭ оказалась однородительская дисомия, т.е. наследование обоих гомологов хромосомы 15 от одного из родителей. С помощью ДНК-маркеров региона делеции путем блотт-гибридизации по Саузерну, а также анализа метилирования было продемонстрировано различное родительское происхождение хромосомы 15:в первом случае отцовское, во втором - материнское. Поскольку этот регион хромосомы 15 идентичен аналогичному региону хромосомы 2мыши, для которого хорошо известен геномный импринтинг, исследования стали проводить в этом направлении. Оказалось, что регион СПВ активен на отцовской хромосоме (при наличии делеции на отцовской хромосоме или материнской ОРД отсутствуют отцовские гены) и не активен на материнской (рис. 3). Из рис. 1 видно, что при СПВ не экспрессируются отцовские гены, а при СЭ - материнские гены. Материнская ОРД наблюдается в 25 % случаев СПВ, а отцовская ОРД становится причиной возникновения СЭ в 3-5 % случаев [1]. В последние годы появились сообщения еще об одной причине развития этих синдромов у пациентов, у которых не было найдено ни типичных делеций, ни ОРД, но зато в семьях таких больных встречались повторные случаи синдрома. В ходе исследования в проксимальном участке хромосомы 15 были обнаружены противоположно импринтированные гены - кандидаты СПВ и СЭ, соответственно SNRPN и UBE3A,в которых выявили мутации. Ген SNRPN кодирует полипептид N малого ядерного рибонуклеопротеина, активно экспрессируется исключительно на отцовской хромосоме 15и репрессирован на материнском гомологе, т.е. мутации в этом гене вовлечены в патогенез СПВ.
Рис. 3 Механизмы возникновения синдромов Прадера-Вилли и Ангельмана через делецию участка хромосомы 15 или однородительскую дисомию по хромосоме 15. СПВ - синдром Прадера-Вилли; СА - синдром Ангельмана; мат. - материнская хромосома; отц. - отцовская хромосома.
Критический регион для СЭ расположен дистальнее (локус D15S10),который экспрессируется только в материнских хромосомах. Предполагают, что мутации при СЭ есть в гене UBE3Aкодирующем убиквитинлигазный белок ЗА. Экспрессия этого белка выявлена во всех тканях человека, причем в ряде структур мозга ген UВЕЗАактивен лишь на материнской хромосоме. Дефицит материнской копии этого гена в клетках Пуркинье (грушевидных невроцитах мозжечка) и нейронах гиппокампа может, по-видимому, объяснить клиническую картину СЭ (умственная отсталость, атаксия, тремор и др.). Таким образом, в районе хромосомы 15(q11-q13)имеются близко расположенные, но противоположно импринтированные локусы, отвечающие за возникновение этих двух синдромов. Эта область хромосомы 15 чрезвычайно существенна для нормальной переустановки геномного импринтинга. Она названа центром импринтинга(IC). Мутации в данной области приводят к ошибкам импринтинга, т.е. теряется способность стирать отпечаток предшествующего поколения. Так, если в сперматогенезе отца не происходит замены «женского» импринта на «мужской» на его материнской хромосоме, то в следующем поколении возникнет состояние, аналогичное материнской ОРД, которое будет сопровождаться фенотипом СПВ. Нарушение установления «женского» эпигенотипа на отцовских хромосомах в овогенезе матери приведет к развитию СА у потомства. Повторный риск для трех групп семей при СПВ и СЭ будет существенно различаться. Так, при делециях он будет ниже 1 %, при ОРД риск также низкий, но в этом случае нужно учитывать возраст матери, который может увеличивать риск. При мутациях в центре импринтинга повторный риск будет существенно выше не только для родителей больного, но и ближайших родственников. Связь геномного импринтинга с другой наследственной патологией человека на уровне хромосом или отдельных генов также отчетливо прослеживается и в настоящее время широко изучается. Так, например, при хорее Гентингтона и спинно-мозжечковой атаксии I заболевание возникает раньше и протекает тяжелее, если унаследованные гены имеют отцовское происхождение. При нейрофиброматозе 1 и 2, миотонической дистрофии, наоборот, заболевание имеет более раннее начало и тяжелое течение при унаследовании мутантных генов от матери. Не вызывает сомнения причастность геномного импринтинга к этиологии опухолевого роста. Выключение импринтинга, а также потеря гетерозиготности или ОРД по хромосомам или их участкам, содержащим импринтированные локусы, могут приводить к функциональной нуллисомии генов-супрессоров опухолевого роста или к аберрантной экспрессии протоонкогенов, что может лежать в основе возникновения рака. Кроме того, вероятность ОРД повышается не только с возрастом матери, но и у носителей изохромосом, робертсоновских и реципрокных транслокаций. Следует иметь в виду, что ОРД (изодисомия) может привести к гомозиготизации определенных регионов хромосомы и быть причиной аутосомно-рецессивной патологии. Такие случаи описаны, например, при муковисцидозе. Точные механизмы, лежащие в основе дифференциальной экспрессии материнских и отцовских геномов, пока не известны. Основную роль в этом процессе отводят специфическому метилированию цитозиновых оснований ДНК. Важнейшими особенностями метилирования ДНК являются, во-первых, стабильное сохранение в ряду многих поколений клеток, а во-вторых, прямое или косвенное влияние на экспрессию генов. Специфическое для пола метилирование некоторых участков генома устанавливается во время гаметогенеза. Известно, что некоторые повторяющиеся и даже уникальные последовательности ДНК являются недометилированными в яйцеклетках и гиперметилированными в сперматозоидах. Такие различия между родительскими хромосомами сохраняются и после оплодотворения и стабильно передаются в следующие клеточные поколения. Как правило, активный ген ассоциируется со сниженным метилированием или его отсутствием, а неэкспрессирующий генетический регион - с гиперметилированием. Тканеспецифичное метилирование цитозиновых остатков ДНК осуществляется в ходе гамето- и эмбриогенеза с помощью ДНК-метилтрансфераз. Значительная доля импринтированных генов (до 15 %) ассоциирует с антисмысловыми транскриптами. Такие транскрипты представлены обычно антисмысловой РНК, происходящей из интронов некоторых генов, и колинеарной ДНК. Эта антисмысловая РНК не выполняет кодирующих функций и, возможно, является регуляторной. Предполагают, что существуют и другие механизмы, регулирующие дифференциальную активность отцовских и материнских генов. Описывают две модели смены эпигенотипа хромосом в гаметогенезе. Согласно первой, переключение эпигенотипа происходит только в той из гомологичных хромосом, которая унаследована от родителя противоположного пола, а вторую хромосому модификации не затрагивают. Вторая модель предполагает предварительное устранение («стирание») существующего эпигенотипа на обеих родительских хромосомах с последующим установлением импринта, соответствующего данному полу. За последние годы в результате многочисленных исследований метилирования и функционирования импринтированных генов в клетках зародышевого пути были получены убедительные доказательства в пользу второго предположения. Таким образом, хотя роль метилирования в обеспечении аллельспецифической экспрессии генов несомненна, остается неясным, является ли метилирование первичным эпигенетическим сигналом, который «стирается» и устанавливается в га-метогенезе, или представляет собой некий вторичный процесс по отношению к более ранней стадии импринтинга и служит лишь для поддержания ранее установленного импринта. Хотя в настоящее время не вызывает сомнения, что метилирование ДНК является эпигенетической меткой и оно достаточно хорошо изучено и характерно практически для всех импринтированных генов и локусов, нельзя исключить и другие пока еще неизвестные механизмы. Дальнейшее изучение геномного импринтинга (особенно в рамках функциональной геномики) будет иметь существенное значение для понимания тонких механизмов регуляции генной активности в онтогенезе и его связи с патологией человека [3]. Импринтинг генов ведет к необычным последствиям. У мужчин материнская копия хромосомы 15 содержит в себе знак того, что она пришла от матери. Но уже в следующем поколении у дочери или сына эта же хромосома будет содержать знак отцовского происхождения. В какой-то момент должно произойти переключение знака хромосомы на противоположный. Нет сомнений в том, что такое переключение происходит, поскольку только этим можно объяснить синдром Ангельмана. Никаких видимых повреждений на хромосоме 15 нет, просто две хромосомы ведут себя так, как будто обе произошли от отца. Это объясняется тем, что в нужный момент в организме матери не произошло переключение знака хромосомы. Возникновение данной проблемы можно проследить в поколениях и обнаружить мутацию в небольшом участке ДНК, непосредственно примыкающем к диверсифицированным генам. Это так называемый центр импринтинга, который каким-то образом указывает на происхождение хромосомы. Импринтинг генов осуществляется с помощью метилирования.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2020-03-26; просмотров: 97; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.12.154.133 (0.009 с.) |