Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Занятие 6. Мембранный потенциал покоя.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Занятие 6. Мембранный потенциал покоя. Потенциал действия Общие свойства возбудимых тканей. Понятие о раздражимости и возбудимости как основе реагирования тканей на раздражение. Опыты Гальвани и Маттеуччи. Возбудимые ткани обладают следующими свойствами: Раздражимость Возбудимость Проводимость Лабильность. 1. Основным свойством живых клеток является раздражимость – это свойство живых клеток отвечать на действие раздражителей изменением процессов их жизнедеятельности, а именно: изменением обмена веществ, теплообразования, роста и размножения клетки. Раздражитель - это фактор внешней или внутренней среды, который действует достаточно сильно, долго и нарастает с достаточной скоростью. Классификация раздражителей. 1. По модальности, т.е. по характеру энергии, свойственной раздражителю: химические (кислоты, щелочи), осмотические физические (тепловые, электрические, световые, звуковые, словесные), биологические (медиаторы, гормоны, микробы); По адекватности Адекватные – это раздражители, которые действуют на данную структуру в естественных условиях и, к восприятию которых она специально приспособлена и чувствительность к ним чрезвычайно велика (например, свет для сетчатки глаза). Неадекватные раздражители - это те раздражители, для восприятия которых данная клетка или орган специально не приспособлены(удар в глаз). По силе действия: Подпороговые Пороговые Надпороговые. 2. Возбудимость – это свойство высокоспециализированных клеток отвечать на действие раздражителя возбуждением. Все ткани реагируют на раздражители, однако условно их делят на возбудимые и невозбудимые. К возбудимым относят нервные, мышечные и некоторые секреторные клетки. Возбуждение — ответ ткани на ее раздражение, проявляющийся в специфической для нее функции (проведение возбуждения нервной тканью, сокращение мышцы, секреция железы) и неспецифических реакциях (генерация потенциала действия, метаболические изменения). 3. Проводимость – способность локального изменения свойств мембраны, возникшего в области действия раздражителя и распространения по протяженности мембраны вплоть до охвата возбуждением всей мембраны клетки. 4. Лабильность – способность ткани ответить на определенное количество стимулов в единицу времени. Строение и основные свойства клеточных мембран и ионных каналов. Согласно современным представлениям биологические мембраны образуют наружную оболочку всех живых клеток. Одним из главных структурных признаков является то, что мембраны всегда образуют замкнутые пространства. Этот факт помогает выполнять им важнейшие функции: 1. Барьерная (создание концентрационных градиентов, что препятствует свободной диффузии веществ). Это обеспечивает создание потенциала покоя, генерацию потенциала действия. 2. Регуляторная (тонкая регуляция внутриклеточного содержимого и внутриклеточных реакций за счет рецепции БАВ, что приводит к изменению активности ферментативных систем мембраны и запуску механизмов вторичных месенджеров (посредников). 3. Преобразование энергии раздражителя в электрические сигналы (в рецепторах). 4. Высвобождение нейромедиаторов в синоптических окончаниях. Химический анализ показал, что мембраны в основном состоят из липидов и белков.
Строение ионного канала.
Различают следующие основные виды электрических ответов возбудимых клеток: 1. Локальные потенциалы (рецепторный, потенциал, локальный ответ, ВПСП (возбуждающий постсинаптический птенциал), ТПСП (тормозной постсинаптический потенциал) Мембранный потенциал покоя (МПП) Потенциал действия (ПД). Мембранный потенциал покоя и его происхождение. Термином «мембранный потенциал покоя » принято называть трансмембранную разность потенциалов, существующую между цитоплазмой и окружающим клетку наружным раствором. Когда клетка (волокно) находится в состоянии физиологического покоя, ее внутренний заряд отрицателен по отношению к наружному, условно принимаемому за нуль. У разных тканей мембранный потенциал характеризуется разной величиной: самый большой у мышечной ткани -80 -90 мВ, у нервной -70 мВ, у соединительной -35 -40 мВ, у эпителиальной -20мВ.
Образование МПП зависит от концентрации ионов К+, Nа+, Са2+, Сl-, и от особенностей строение мембраны клетки. В частности важным в образовании МПП является то, что мембранные ионные каналы, обладают свойствами: 1. Селективностью (избирательной проницаемостью) 2. Электровозбудимостью. В состоянии покоя натриевые каналы все закрыты, а большинство калиевых – открыты. Каналы могут открываться и закрываться. В мембране существуют каналы утечки (неспецифические), которые проницаемы для всех элементов, но более проницаемы для калия. Ионы движутся через эти каналы по концентрационному и электрохимическому градиенту. Согласно мембранно-ионной теории наличие МПП обусловлено: Ø непрерывным движением ионов по ионным каналам мембраны, Ø постоянно существующей разностью концентраций катионов по обе стороны мембраны, Ø непрерывной работой натрий-калиевого насоса. Ø различной проницаемостью каналов для этих ионов. В состоянии покоя мембрана клетки проницаема только для ионов К+. Ионы калия в состоянии покоя постоянно выходят в окружающую среду, где высокая концентрация Nа+. Поэтому, в состоянии покоя, наружная поверхность мембраны заряжена положительно. Высокомолекулярные органические анионы (белки) концентрируются у внутренней поверхности мембраны и определяют ее отрицательный заряд. Они же электростатически удерживают ионы К+ с другой стороны мембраны. Основную роль в образовании МПП принадлежит ионам К+. Блокируют проницаемость К+ тетраэтиламмония, аминопиридины. Несмотря на потоки ионов через каналы утечки разность концентрации ионов не выравнивается, т.е. сохраняется всегда постоянной. Этого не происходит потому, что в мембранах существуют Nа+ - К+- насосы. Они непрерывно откачивают Nа+ из клетки и против градиента концентрации вводят в цитоплазму К+. На 3 иона Nа+, которые выводятся из клетки, внутрь вводится 2 иона К+. Перенос ионов против градиента концентрации осуществляется активным транспортом (с затратой энергии). В случае отсутствия энергии АТФ клетка погибает. Наличие потенциала покоя позволяет клетке практически мгновенно после действия раздражителя перейти из состояния функционального покоя в состояние возбуждения. При возбуждении происходит снижение величины исходного потенциала покоя с перезарядкой мембраны. Когда внутренний заряд мембраны становится менее отрицательным наступает деполяризация мембраны и начинает развиваться потенциал действия, увеличение отрицательного – гиперполяризация (возбудимость снижается). Потенциал действия и механизм его происхождения. Соотношение фаз возбудимости с фазами потенциала действия Потенциалом действия называют быстрое колебание мембранного потенциала, возникающее при действии раздражителя пороговой силы в нервных и мышечных клетках. В его основе лежат изменения ионной проницаемости мембраны. Амплитуда и характер изменений потенциала действия мало зависят от силы вызывающего его раздражителя, важно лишь, чтобы эта сила была не меньше пороговой. Возникнув, ПД распространяется вдоль мембраны, не изменяя своей амплитуды. В нем различают фазы: 1) Деполяризации: а) медленная деполяризация; б) быстрая деполяризация. 2) Реполяризация: а) быстрая реполяризация; б) медленная реполяризация (отрицательный следовой потенциал) 3) Гиперполяризация (положительный следовой потенциал)
Возникновение потенциала действия (деполяризация)
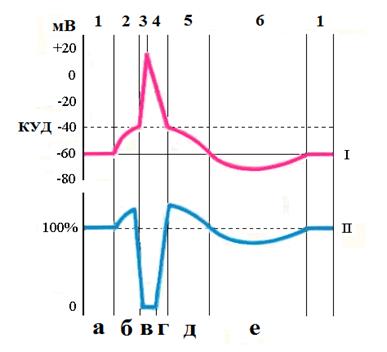
I – мембранный потенциал 1 - потенциал покоя, 2-- медленная деполяризация, 3 - быстрая деполяризация, 4 - быстрая реполяризация, 5 – медленная реполяризация, 6- гиперполяризация II - возбудимость а – нормальная, б – повышенная, в –абсолютная рефрактерность, г – относительная рефрактерность, д – супернормальность, е -субнормальность Механизм возникновения ПД. Деполяризация После действия раздражителя пороговой силы потенциал покоя уменьшается, повышает проницаемость каналов для ионов натрия, который входит в клетку. Поток ионов натрия в клетку выше, чем поток ионов К+ из клетки. Поступление в клетку иона натрия уменьшает электроотрицательность внутренней поверхности мембраны, что способствует активации новых ионных натриевых каналов и дальнейшему поступлению в клетку иона натрия. Внутренняя сторона мембраны становится положительной, а на поверхности ионы Cl- образуют отрицательный заряд. Перезарядка мембраны, или овершут, характерен для большинства возбудимых клеток. Амплитуда овершута характеризует состояние мембраны и зависит от состава вне- и внутриклеточной среды. Смена зарядов на внутренней и наружной поверхности мембраны соответствует фазе деполяризации. Реполяризация После смены зарядов на наружней и внутренней поверхности клеточной мембраны происходит инактивации (закрытия) натриевых каналов. Поступление в клетку ионов натрия становится минимальным. Выход из клетки ионов калия восстанавливает электроотрицательность внутренней поверхности мембраны, что соответствует фазе реполяризации. В последующем натрий/калиевый насос мембраны удаляет из клетки поступивший при деполяризации натрий и восстанавливает исходную концентрацию калия, который вышел из клетки при реполяризации. Основная роль в образовании ПД принадлежит ионам Na+. Блокирует транспорт нартия тетродотоксин. Лабильность. Для характеристики протекания отдельных ПД используется понятие лабильность. Лабильность – это скорость развития ответа на раздражитель (отдельных ПД). Чем выше лабильность тем больше ПД может произвести ткань в единицу времени. Мерой лабильности является наибольшее количество импульсов, которое ткань может генерировать в единицу времени. Максимальный ритм возбуждения лимитируется длительностью периода абсолютной рефрактерности. Если рефрактерность длиться 0,5 мс, то максимальный ритм составляет 1000 импульсов в секунду и выше. Самой высокой лабильностью обладает нервная ткань. Она способна генерировать до 1000 импульсов в секунду. Мышечная ткань способна проводить до 500 импульсов в секунду. Наименьшей лабильность обладают синапсы. При этом в максимальном ритме ткань не может функционировать долго. В естественных условиях ткани реагируют на возбуждение в более низком ритме, который может сохраняться длительный период времени. Формируется этот ритм через период супернормальности и поэтому называется оптимальным. Так, у нервного волокна он составляет 500 импульсов в секунду, у мышцы 200 импульсов в секунду. В ходе ритмического возбуждения лабильность может увеличиваться или уменьшаться. Снижение лабильности ведет к развитию процессов торможения, а ее увеличение определяет свойства ткани усваивать новые более высокие ритмы импульсации. Усвоение более высокого ритма связано с выкачиванием ионов Nа+ из цитоплазмы, когда возбуждение проникает внутрь клетки. Таким образом мышцы способны усваивать более частый ритм импульсов поступающих к ним от нервных волокон. В практической медицине используют эти фундаментальные знания — Местные анестетики блокируют натриевые каналы. Натрий не входит в нервные волокна, значит возбуждение не возникает (боль не распространяется). — Кардиологи управляют входом Кальция (лечение аритмий, гипертонии) Схема нервного ствола. ФУНКЦИИ НЕРВОВ — Информационная - передача ПД от тела нейрона на периферию и обратно. — Трофическая – трафик нутриентов, органелл. — Генетическая детерминация направления роста. Классификация нервных волокон. 1. По строению нервы делятся на: Ø миелинизированные (мякотные) Ø немиелинизированные. Миелиновые волокна имеют прерывистую оболочку, которая образована сегментами миелина длиной 1—2 мм. Участок между двумя сегментами называется перехватом Ранвье. На поверхности миелин покрыт Швановскими клетками. Миелин обладает высоким сопротивлением и поэтому выполняет изоляционную функцию и кроме того он принимает участие в обмене веществ нервного волокна. Возбуждение по миелиновому волокну распространяется только через перехваты Ранвье так как там много натриевых каналов. Безмиелиновые волокна обладают такой же структурой, но не имеют миелина. их поверхность покрыта Швановской клеткой. Если перерезать нервное волокно, то его периферический конец через некоторое время теряет способность проводить возбуждение и дегенерирует. Перерождение периферического конца нервного волокна изучал Валлер (в 19 столетии) поэтому назвали Валлеровское перерождение. Миелин претерпевает жировое перерождение и превращается в жировые капли. Центральный конец нервного волокна способен регенерировать. На нем образуется колба роста, которая растет к периферии (в сутки от 0,4 до 4,5 мм) и достигает соответствующего органа или ткани. Таким образом, восстанавливается их иннервация. Так, первые признаки восстановления иннервации мышц могут появляться через 5—6 недель. 2. По направлению передачи информации (центр - периферия) нервы подразделяются на: Ø афферентные Ø эфферентные. Эфферентные по физиологическому эффекту делятся на: Ø Двигательные (иннервируют мышцы). Ø Сосудодвигательные (иннервируют сосуды). Ø Секреторные (иннервируют железы). 3. По характеру влияния на эффекторный орган нейроны делятся на: Ø пусковые (переводят ткань из состояния физиологического покоя в состояние активности) Ø корригирующие (изменяют активность функционирующего органа). 4. По функциональным свойствам (строению, диаметру волокна, электровозбудимости, скорости развития потенциала действия, длительности различных фаз потенциала действия, по скорости проведения возбуждения) нервные волокна разделили на группы А, В и С. Группа А неоднородна. Волокна этого типа в свою очередь делятся на подтипы: А-альфа, А-бета, А-гамма, А-дельта. А-альфа – самые толстые волокна покрыты миелиновой оболочкой. Их диаметр от 12 до 22 мкм. Скорость распространения возбуждения по такому волокну составляет от 70 до 120 мс, ПД самый короткий. Эти волокна проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам (двигательные волокна) и от проприорецепторов мышц к соответствующим нервным центрам. Три другие группы волокон типа А (бета, гамма, дельта) имеют меньший диаметр от 8 до 1 мкм и меньшую скорость проведения возбуждения от 5 до 70 м/с. Волокна этих групп относятся преимущественно к чувствительным, проводящим возбуждение от различных рецепторов (тактильных, температурных, некоторых болевых рецепторов внутренних органов) в ЦНС. Исключение составляют лишь гамма-волокна, значительная часть которых проводит возбуждение от клеток спинного мозга к интрафузальным мышечным волокнам. А-бета – диаметр от 8 до 13 мкм. Скорость распространения возбуждения от 40 до 70 мс. А-гамма – диаметр от 4 до 8 мкм. Скорость распространения возбуждения от 15 до 40 мс. А-дельта – диаметр от 1 до 4 мкм. Скорость распространения возбуждения от 5 до 15 мс. К волокнам типа В относятся миелинизированные преганглионарные волокна вегетативной нервной системы. Их диаметр — 1- 3 мкм, а скорость проведения возбуждения — 3-18 м/с, ПД длиться 1—2 мс. У эти волокон отсутствует период супернормальности. К волокнам типа С относятся безмиелиновые нервные волокна малого диаметра — 0,5-2,0 мкм. Скорость проведения возбуждения в этих волокнах не более от 0,5 до 3 м/с, ПД длиться 2 мс. Большинство волокон типа С — это постганглионарные волокна симпатического отдела вегетативной нервной системы, а также нервные волокна, которые проводят возбуждение от болевых рецепторов, некоторых терморецепторов и рецепторов давления. Таким образом, скорость проведения возбуждения по нерву пропорциональна его диаметру. Нервы практически не утомляются. Они способны очень длительное время проводить импульсы. Механизмы проведения нервного импульса в Классификация синапсов. 1. По местоположению и принадлежности соответствующим структурам: Ø периферические (нервно-мышечные, нейросекреторные, рецепторнонейрональные); Ø центральные (аксо-соматические, аксо-дендритные, аксо-аксональные, сомато-дендритные, сомато-соматические); 2. По эффекту действия: Ø возбуждающие Ø тормозные 3. По способу передачи сигналов: Ø электрические. Передают возбуждение без участия медиатора с большой скоростью и обладают двухсторонним проведением возбуждения. Структурной основой электрического синапса является нексус. Встречаются эти синапсы в железах внутренней секреции, эпителиальной ткани, ЦНС, сердце. Ø химические, Ø смешанные. В некоторых органах возбуждение может передаваться и через химические и через электрические синапсы. 4) По медиатору, с помощью которого осуществляется передача сигнала химические синапсы классифицируют: Ø холинергические, Ø адренергические, Ø серотонинергические, Ø глицинергически. и т.д. Синапсы с химической передачей возбуждения обладают рядом общих свойств: Ø Возбуждение через синапсы проводится только в одном направлении (односторонне). Это обусловлено строением синапса: медиатор выделяется только из пресинаптического утолщения и взаимодействует с рецепторами постсинаптической мембраны; Ø передача возбуждения через синапсы осуществляется медленнее, чем по нервному волокну — синаптическая задержка; Ø передача возбуждения осуществляется с помощью специальных химических посредников — медиаторов; Ø в синапсах происходит трансформация ритма возбуждения; Ø синапсы обладают низкой лабильностью; Ø синапсы обладают высокой утомляемостью; Ø синапсы обладают высокой чувствительностью к химическим (в том числе и к фармакологическим: блокаторам, психомиметикам). Занятие 6. Мембранный потенциал покоя. Потенциал действия
|
||||
|
|
Последнее изменение этой страницы: 2019-08-19; просмотров: 995; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.20 (0.016 с.) |