
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Дайте определение растительным тканям. Общая характеристика и принципы классификации. Типы тканей.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Дайте определение растительным тканям. Общая характеристика и принципы классификации. Типы тканей. Ткани появились у высших растений в связи со специализацией клеток. Ткань – совокупность клеток и межклеточного вещества, сходных по происхождению, строению и выполняемым функциям. Различают простые и сложные ткани. Если ткань состоит из одинаковых клеток, как например, паренхима, то это простая ткань. Сложные ткани имеют общее происхождение, и выполняют единую функцию, но различные клетки сложной ткани сильно отличаются друг от друга. Например, древесина (ксилема) – сложная ткань, в состав которой входит проводящая (трахеи и трахеиды), механическая (древесные волокна) и основная (древесная паренхима) ткани. Существует много классификаций тканей. Например: мёртвые и живые, паренхимные и прозенхимные (соотношение длинны и ширины клетки) -Паренхимные клетки имеют примерно одинаковые длину и ширину, т.е. эти клетки изодиаметрические. Их форма наиболее разнообразна: округлая, овальная многогранная, дисковидная, таблитчатая, звездчатая и др., как правило, живые и тонкостенные. Они составляют основные ткани растения – сердцевину и кору стебля, корня, ткани листа, цветка и семени, мякоть плодов. -Прозенхимные клетки вытянутые, длина их превышает ширину в десятки и сотни раз. Окончания клеток заостренные, оболочки толстые, содержимое часто отсутствует. Прозенхимные клетки образуют главным образом проводящие и механические ткани растений. Живые физиологически активные клетки редко имеют прозенхимную форму. Общепринятая классификация тканей растений исходит из их функций и анатомического строения. В соответствии с этим все ткани делятся на 2 группы: 1. Образовательные ткани или меристемы; Дайте характеристику строение семян и проростков. Надземное и подземное прорастание. Стратификация и скарификация.
Строение семян голосеменных Семя развивается на поверхности семенной чешуи. Оно представляет собой многоклеточную структуру, объединяющую запасающую ткань — эндосперм, зародыш и специальный защитный покров (семенную кожуру). До оплодотворения в центральной части семязачатка имеется нуцеллус, который постепенно вытесняется эндоспермом. Эндосперм гаплоидный и образуется из тканей женского гаметофита.
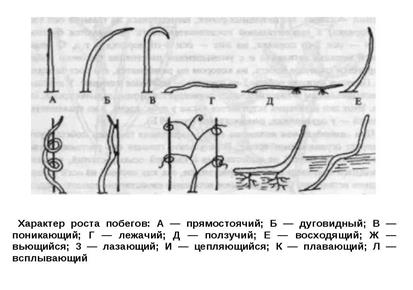
Определение в етвления, его типы и биологическое значение. Охарактеризуйте листорасположение и направление роста побегов. Строение и деятельность конуса нарастания. Специализация и метаморфоз побегов. Ветвление — процесс образования новых побегов и характер их взаимного расположения на стебле, многолетней ветви и корневище. Поскольку побег является осевым органом, у него присутствует апикальная меристема, которая обеспечивает неограниченное нарастание в длину. Такой рост сопровождается более или менее регулярным ветвлением побега. У низших растений в результате ветвления возникает разветвлённый таллом (слоевище), у высших— образуются системы побегов и корней. Ветвление позволяет многократно увеличивать фотосинтетическую поверхность и обеспечивать растение органическими веществами. У различных растений наблюдается несколько типов ветвления: дихотомическое, моноподиальное, симподиальное. При дихотомическом ветвлении расщепляется (раздваивается) зона роста, находящаяся на верхушке каждой оси. Поэтому дихотомическое ветвление называют также верхушечным. Исходной для эволюции этого ветвления была равная дихотомия — изотомия (рис. 13 А), при которой обе веточки росли с одинаковой скоростью, а затем их верхушки снова раздваивались. Если одна из веточек опережала в развитии другую, возникала неравная дихотомия — анизотомия (рис. 13 Б). Резкое отставание развития одной из веточек при водило к дихоподиальному ветвлению (рис. 13В), при этом формировалась зигзагообразно изогнутая главная ось растения. Из дихотомического ветвления развились 2 типа боковых ветвлений. (Дихотомическое ветвление — это наиболее древний тип ветвления, оно свойственно низшим растениям, а среди высших растений встречается у мхов, плаунов. При дихотомическом ветвлении верхушечная почка главного побега разделяется в процессе формирования на две почки, которые и дают два новых побега следующего порядка, те, в свою очередь, таким же путем дают побеги третьего порядка и т.д.)
При моноподиальном ветвлении верхушечная почка (конус нарастания) функционирует в течение всей жизни растения, образуя главный побег (у древесных растений — ствол). Из пазушных почек, находящихся на главной оси (оси 1-го порядка) в акропетальной последовательности развиваются боковые побеги — оси 2-го порядка, на них — оси 3-го порядка и т.д. С возрастанием порядка ветвления и с уменьшением расстояния бокового побега от верхушки главного побега, на котором он развивается, его рост замедляется. Поэтому нижние побеги 2-го порядка — наиболее длинные. У деревьев с моноподиальным ветвлением часто формируется пирамидальная крона, как у ели. Этот тип ветвления встречается также у дуба, клена, а из травянистых растений — у одуванчика, подорожника (При моноподиальном ветвлении главный стебель растёт за счёт верхушечной почки в течение всей жизни растения. На главном побеге за счёт пазушных почек развиваются боковые побеги (оси второго порядка), на них развиваются оси третьего порядка и т.д. Главный побег растёт и утолщается сильнее боковых, растение обычно имеет стройную, вытянутую форму.)
При симподиальном ветвлении конус нарастания главного побега функционирует ограниченное время. В результате главная ось растения (ствол у древесных растений) не сопоставима с главной осью растений, характеризующихся моноподиальным нарастанием, так как состоит из осей разных порядков ветвления. Симподиальное ветвление свойственно тополю, березе, липе, багульнику, бруснике и другим растениям Разновидность симподиального ветвления, называемая псевдо-дихотомическим (ложно-вильчатым), нередко встречается у растений с супротивными листьями — у омелы, свидины, сирени. У них из верхушечной почки развивается генеративный побег — одиночный цветок или соцветие, и дальнейший рост главного побега прекращается (Симподиальное ветвление характеризуется тем, что верхушечная почка главного и боковых побегов спустя некоторое время отмирает и вместо неё начинает развиваться ближайшая к верхушке боковая почка, которая образует побег, растущий вертикально. Таким образом, при симподиальном ветвлении побеги состоят из осей разных порядков, сменяющих друг друга. В результате этого образуется сильно раскидистая крона, так как отмирание верхушечных почек стимулирует заложение и развитие боковых.)
Ветвление имеет важное значение в жизни растения, увеличивая ассимилирующую поверхность и улучшая, таким образом, его питание. Вместе с этим возрастает и общее количество меристемы, слагающей конусы нарастания побегов еще не развернувшихся почек. Так как меристема реализуется не сразу, растение всегда имеет «запас» этой ткани, используемый не только на естественное увеличение мощности побеговой системы, но и на восстановительные процессы после какого-либо повреждения.
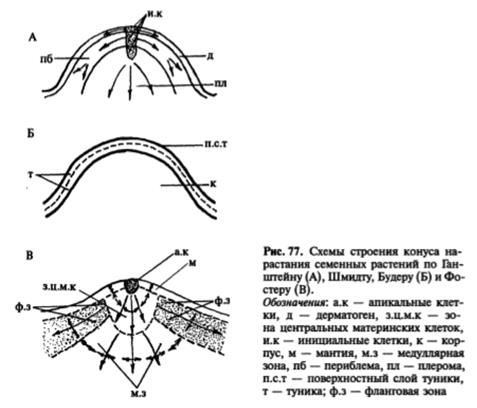
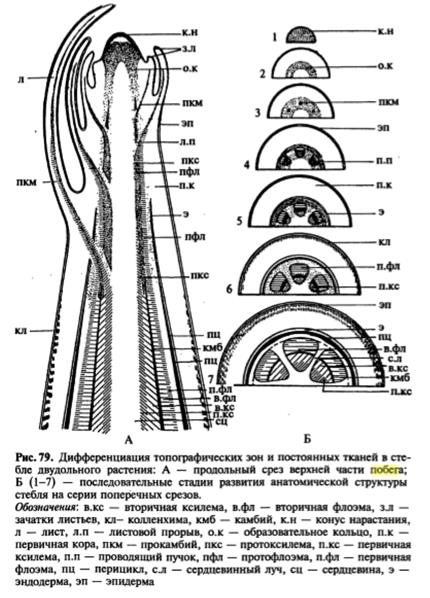
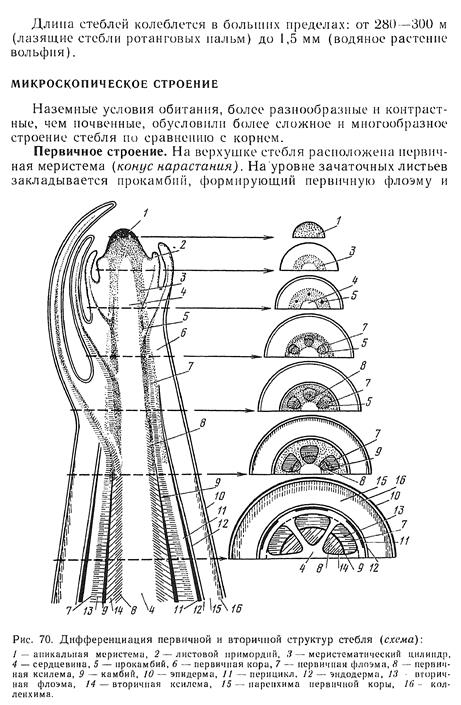
Строение конуса нарастания. Конусы нарастания побегов голосеменных и покрытосеменных растений отличаются от таковых высших споровых отсутствием четко выраженных инициалей и тенденцией к расположению клеток апикальной меристемы несколькими слоями. В 1868 г. немецкий ботаник Дж. Ганштейн выявил в конусе нарастания покрытосеменных три слоя клеток, названных им гистогенами (от греч. histos — ткань и genos — род, происхождение): дерматоген — самый поверхностный слой, дающий начало эпидерме; 1-5-слойная периблема, образующая ткани первичной коры; многослойная плерома, из которой развиваются ткани центрального цилиндра, или стелы.
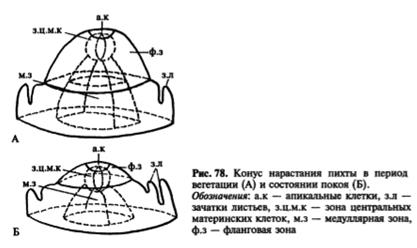
Верхнюю часть конуса нарастания составля.ет группа апикальных клеток, участвующих в образовании поверхностного слоя — мантии и корпуса, в котором различимы три зоны: зона центральных материнских клеток, находящаяся под апикальными клетками, расположенная под ней медуллярная зона и фланговая зона, прилегающая к мантии (рис. 77 В). Центральные материнские клетки самые крупные в конусе нарастания. Они изодиаметрические, сильно вакуолизированные. Эти клетки участвуют в образовании медуллярной и фланговой зон. Клетки медуллярной зоны нередко расположены продольными рядами, эту зону называют также стержневой. Она образует сердцевину. Фланговая зона состоит из типичной эвмеристемы, для которой характерны мелкие, изодиаметрические, тонкостенные, крупноядерные клетки. В отличие от остальных клеток конуса нарастания клетки фланговой зоны делятся регулярно, участвуя в образовании листовых бугорков, прокамбия и части первичной коры. Размеры и форма конуса нарастания побега не постоянны даже у одного и того же растения. Они подвержены возрастным и сезонным изменениям. Наиболее изменчивы размеры фланговой и медуллярной зон, которые увеличиваются весной и летом, в период активной деятельности конуса нарастания, образующего метамеры побега. Осенью и зимой размеры этих зон значительно меньше. Более постоянны размеры мантии и зоны центральных материнских клеток. В период заложения листовых бугорков конус нарастания немного уплощается, после образования листового бугорка он вытягивается.
Зональность — не изначальное свойство конуса нарастания, в разные периоды жизни растения она выражена по разному. У проростков конус нарастания имеет только мантию и зону центральных материнских клеток. В вегетативной фазе развития растения, когда происходит образование метамеров побега, в конусе нарастания хорошо выражены все зоны, наиболее активную роль играет фланговая, участвующая в заложении зачатков листьев. Внутри почки заключена меристематическая верхушка побега - его апекс, основная часть которого представляет собой конус нарастания. Деятельность конуса нарастания обеспечивает формирование всех тканей и частей побега, т.е. его органогенез и гистогенез. Конус нарастания не обязательно имеет геометрическую форму конуса, нередко это плоская или даже вогнутая вершинная часть апекса, сложенная особыми клетками - инициалями верхушечной меристемы и ближайшими их производными. В результате меристематической активности апекса образуются зачатки листьев (листовые примордии), зачаточные узлы и междоузлия. В пазухах зачаточных листьев обычно располагаются зачатки почек следующего порядка. Ритм появления зачаточных листьев характерен для каждого растения и в известной мере определяет характер листорасположения взрослого побега.
Меристематическая активность конуса нарастания в разных его частях неодинакова. Существует ряд теорий, объясняющих неодинаковый характер деятельности его разных зон, в частности, теория "туники-корпуса", сформулированная немецким ботаником А.Шмидтом. Согласно этой теории конус нарастания у покрытосеменных и части голосеменных состоит из двух гистологически разных частей, обладающих различной меристематической активностью. Наружная часть конуса нарастания получила название туники, а внутренняя - корпуса. Самые наружные клетки туники - клетки протодермы - образуют первичную покровную ткань - эпидерму, а лежащие глубже - внутренние ткани листа и первичную кору стебля. Клетки корпуса делятся во всех направлениях и обеспечивают объемный рост апекса. На границе туники и корпуса на уровне листовых зачатков закладывается так называемая периферическая меристема, или инициальное кольцо. Именно здесь в этой меристеме закладываются тяжи относительно узких и длинных, продольно делящихся клеток прокамбия, дающих начало проводящим пучкам. Прокамбий распространяется по мере дальнейшего роста и в глубь растущего листового зачатка, и в глубь оформляющегося стебля - оси побега, образуя основу будущей проводящей системы побега, связывающей листья и стебли.
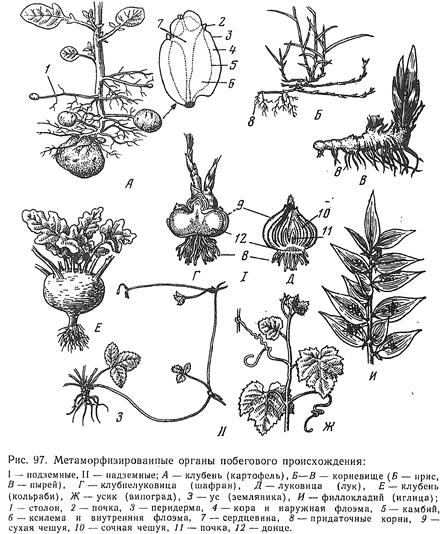
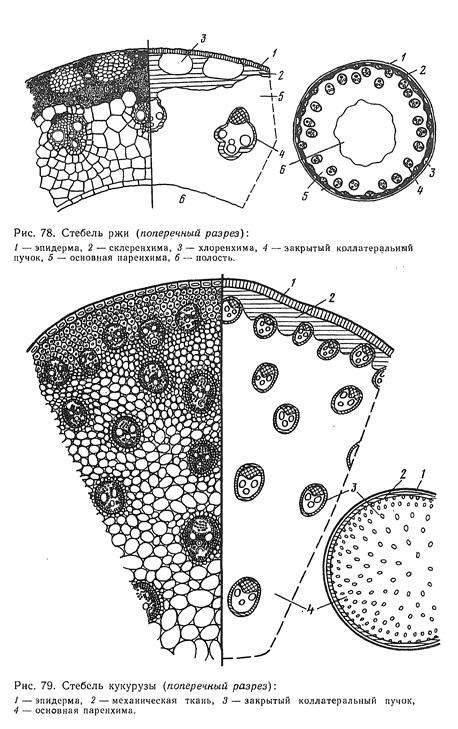
Наиболее обычный случай специализации представляют собой ствол и сучья деревьев и кустарников. Известно, что их побеги через один (у листопадных) или несколько лет (у вечнозеленых) теряют ассимиляционную функцию. Часть таких побегов иногда отмирает, но большинство остается, выполняя опорную, проводящую и запасающую функции. Из них формируется система скелетных осей, утолщающихся за счет деятельности камбия. Лишенные листьев скелетные оси известны под названием сучьев, стволов (у деревьев) и стволиков (у кустарников). Дайте характеристику стеблю, его функции и особенности морфологии. Первичное строение стебля. Переход ко вторичному утолщению и работа камбия. Сравнительно-анатомическая характеристика травянистых двудольных и однодольных растений.
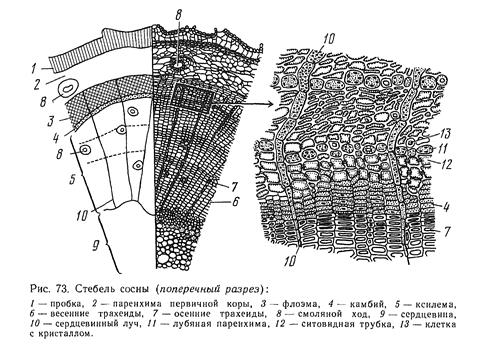
5. Опишите анатомическое строение стебля покрытосеменных (на примере липы) и голосеменных растений (на примере сосны). Приведите примеры.
СИСТЕМАТИКА ВОДОРОСЛЕЙ В систематическом отношении водоросли делятся на множество самостоятельных отделов, различающихся по окраске, зависящей от набора пигментов, по организации клетки и структуре талломов. Выделяют следующие отделы: прокариотические - Cyanophyta – Сине-зелёные; эукариотические - Chlorophyta – Зелёные; Euglenophyta – Эвгленовые; Chrysophyta – Золотистые; Dinophyta – Динофитовые; Xanthophyta – Желто-зелёные; Bacillariophyta (Diatomophyta) – Диатомовые; Phaeophyta – Бурые; Rhodophyta – Красные.
Экология водорослей Планктонные водоросли Фитопланктоном называют совокупность свободноплавающих (в толще воды) мелких, преимущественно микроскопических, растений, основную массу которых составляют водоросли. Соответственно каждый отдельный организм из состава фитопланктона называют фитопланктером.
Фитопланктон существует в водоемах самой разной природы и самых разных размеров — от океана до маленькой лужи. Его нет лишь в водоемах с резко аномальным режимом, в том числе в термальных (при температуре воды выше +70, +80 °С), заморных (зараженных сероводородом), чистых приледниковых водах, не содержащих минеральных питательных веществ. Морской фитопланктон состоит в основном из диатомовых и перидиниевых водорослей. Характерной морфологической особенностью представителей морского фитопланктона является образование у них разного рода выростов: щетинок и острых шипов у диатомовых, воротничков, лопастей и парашютов у перидиней. Похожие образования встречаются и у пресноводных видов, но там они выражены значительно слабее. Пресноводный фитопланктон отличается от типично морского огромным разнообразием зеленых и сине-зеленых водорослей. Особенно многочисленны среди зеленых одноклеточные и колониальные вольвоксовые и протококковые. Бентосные водоросли К числу бентосным (донных) водорослей относятся водоросли, приспособленные к существованию в прикрепленном состоянии на дне водоемов и на разнообразных предметах, живых и мертвых организмах, находящихся в воде. В зависимости от места произрастания среди бентосных водорослей различаются: 1) эпилеты, которые растут на поверхности твердого грунта (скалы, камни и т. д.); 2) эпипелеты, населяющие поверхность рыхлых грунтов (песок, ил); 3) эпифиты, живущие на поверхности других водорослей; 4) эндолиты, или сверлящие водоросли, внедряющиеся в известковый субстрат (скалы, раковины моллюсков, панцири ракообразных); 5) эндофиты, которые поселяются в слоевищах других водорослей, но, в отличие от паразитических видов, содержат нормальные хлоропласта в клетках; 6) паразиты, живущие в слоевищах других водорослей, хлоропласты в клетках не выражены. Иногда водоросли, растущие на предметах, введенных в воду человеком (суда, плоты, буи), относят к перифитону. Выделение этой группы обосновывают тем, что входящие в ее состав организмы (водоросли и животные) живут на предметах, большей частью находящихся в движении или обтекаемых водой. Наземные водоросли Экологическую группировку наземных водорослей составляют все те формы, которые обитают вне водоемов на поверхности различных твердых субстратов, благодаря, чему они в течение всей жизни окружены воздухом. Эти водоросли поселяются на стволах деревьев и кустарников, на мхах, валунах и скалах, во влажном тропическом климате — на листьях деревьев и кустарников и надземных частях травянистых растениях, а также на заборах, стенках домов, крышках и т.п. (диатомовые, сине-зеленые водоросли) Почвенные водоросли На поверхности почвы нередко можно видеть невооруженным глазом различные разрастания — кожистые или войлокообразные пленки или слизистые слоевища сине-зеленых водорослей. Часто наблюдается также общее позеленение почвы, обусловленное массовым развитием микроскопических форм, рассеянных среди почвенных частиц. Общее количество обнаруженных в почве видов водорослей приближается уже к 2000. Относящихся главным образом к сине-зеленым, зеленым, желто-зеленым и диатомовым водорослям. Почва как среда обитания характеризуется целым рядом экологических особенностей. Она сходна как с водными, так и с воздушными местообитаниями: в почве есть воздух, но насыщенный водяными парами, что обеспечивает дыхание атмосферным воздухом без угрозы высыхания Почвенные водоросли отличаются изменчивостью способа питания. На небольших глубинах, в пределах проникновения света, они, как и высшие растения, являются типичными фототрофами. В более глубокие горизонты водоросли заносятся с поверхности путем вмывания, а также почвенными животными, и корнями растений. Поэтому в глубоких слоях почвы обнаруживается сравнительно небольшое число видов, преимущественно одноклеточные зеленые и желто-зеленые водоросли. Особенность почвенных водорослей - «эфемерность» их вегетации, т. е. способность быстро переходить из состояния покоя к активной жизни и наоборот. Во многих случаях у водорослей, обитающих на поверхности почвы, появляются приспособления для защиты от избыточного освещения — темные слизистые чехлы вокруг клеток. Особенно устойчивы против ультрафиолетового облучения сине-зеленые водоросли. Почвенные водоросли обладают устойчивостью против радиоактивных излучений. Благодаря перечисленным приспособлениям почвенные водоросли способны существовать даже при крайне неблагоприятных условиях среды. Этим объясняются широкое распространение почвенных и наземных водорослей и быстрота их разрастания даже при кратковременном появлении необходимых факторов. Почвенные водоросли представляют большой общебиологический интерес как организмы необыкновенной выносливости и устойчивости к крайним условиям существования. Водоросли снега и льда Прямую противоположность теплолюбивым (термофильным) водорослям составляет группировка развивающихся на поверхности снега и льда холодолюбивых, или криофильных, водорослей. В этих, казалось бы, крайне неблагоприятных условиях могут жить многие водоросли, причем они размножаются здесь столь интенсивно, что своей массой явственно окрашивают поверхность снега и льда. Наибольшую известность с давних пор получило явление так называемого «красного снега». Главным организмом, вызывающим окраску снега, является один из видов хламидомонады, названный хламидомонадой снежной. Большую часть времени эта Все же чаще других встречается зеленое «цветение» снега, вызываемое различными видами зеленых водорослей. Не менее интенсивное развитие водорослей наблюдается и во льдах арктических и антарктических бассейнов. Это — подлинная стихия диатомей, размножающихся здесь в огромных количествах и окрашивающих лед в грязно-бурый или желто-коричневый цвет. Все эти водоросли приспособлены к жизни в крайне неблагоприятных условиях низких температур. Находясь в поверхностных слоях снега и льда, они подвергаются очень сильному охлаждению в зимние стужи, когда температура воздуха опускается на несколько десятков градусов ниже нуля, а в летнее время живут и размножаются в талой воде, т. е. при температуре около 0°С. И если снежная хламидомонада имеет стадию покоя в виде округлых толстостенных клеток, то многие другие водоросли, в том числе и диатомей, лишены каких-либо специальных приспособлений для перенесения столь низких температур. Водоросли соленых водоемов К числу факторов, создающих особые условия для жизни водорослей, относится также повышенное содержание в воде солей, свойственное некоторым связанным с морем и континентальным водоемам. Количество видов водорослей по мере увеличения солености убывает, очень высокую соленость переносят только немногие из них, но в целом солевыносливых форм немало. Из зеленых водорослей в водоемах с большой концентрацией солей (до 285 г на литр) широко распространена и чрезвычайно характерна дюналиелла, микроскопическая одноклеточная подвижная водоросль из порядка вольвоксовых. Тело дюналиеллы грушевидной или яйцевидной формы, заостренное на переднем конце, где располагаются два жгута. Явственной, отделимой от протопласта оболочки нет — только наружная уплотненная пленка. Содержимое клетки такое же, как у хламидомонад; кроме того, имеется еще красный пигмент гематохром, маскирующий зеленый цвет хлоропласта. При массовом размножении, когда клетки дюналиеллы отмирают, ее пигменты сообщают солевому раствору (рапе) и выпадающей из него соли в пересолоненных водоемах характерную окраску — от розовой до красной. Из сине-зеленых водорослей большой интерес представляет хлороглея сарциноидная, в огромном количестве развивающаяся в некоторых солевых озерах с высокой концентрацией солей. Приспособившись к столь необычным условиям существования, эти водоросли играют очень большую роль в жизни соленых водоемов. Сочетание органической массы, образуемой водорослями, и большого количества растворенных в воде солей обусловливает ряд своеобразных биохимических процессов, свойственных этим водоемам. В частности, хлороглея и ряд других водорослей, тоже размножающихся в массовом количестве, участвуют в некоторых озерах (например, в Мойнакском) в процессе бразования лечебных грязей. ГРИБЫ Общая характеристика грибов Наука о строении, жизнедеятельности, особенности циклов развития, экологии, географии и значении грибов называется микология. Предмет изучения – высшие и низшие грибы. Грибы – это обширная группа эукариотических гетеротрофных организмов, насчитывающая более 100 тыс. видов, занимающая особое положение в системе органического мира. Клетка грибов имеет эукариотическое строение. В большинстве случаев покрыта твёрдой оболочкой – клеточной стенкой. У большинства грибов основными полисахаридами, входящими в состав клеточных стенок, являются хитин и хитозан (у оомицетов – целлюлоза). У молодых клеток грибов оболочка тонкая, бесструктурная, бесцветная. С возрастом клетки, внутри образовываются новые слои оболочки, она темнеет за счёт пигмента меланина. Ядра мелкие, сферические, либо овальные. В клетке грибов присутствуют вакуоли, содержащие запасные питательные вещества – волютины, липиды, гликоген, жирные кислоты. У настоящих грибов имеются признаки, сближающие их с растениями: образование хорошо выраженной клеточной стенки; осмотрофное питание; размножение спорами; неподвижность в вегетативном состоянии; неограниченный рост; а также признаки, сближающие грибы с животными: первично гетеротрофный способ питания; наличие в клеточной стенке хитина; запасной продукт – гликоген; выделение продукта жизнедеятельности – мочевины; отсутствие пластид и неспособность к фотосинтезу; путь синтеза лизина через аминоадипиновую кислоту (у растений – через диаминопимелиновую кислоту). Кроме того, имеются и специфические признаки: способность к синтезу маннитола (за исключением зигомикотовых грибов), собственные ферментативные системы синтеза триптофана, отсутствие комплекса Гольджи и его замена видоизменениями эндоплазматического ретикулума. Вегетативное тело большинства грибов представляет собой мицелий, состоящий из ветвящихся нитей – гиф с апикальным (верхушечным) ростом и боковым ветвлением. Мицелий пронизывает субстрат и всей поверхностью поглощает из него питательные вещества (субстратный мицелий), а также располагается на его поверхности и может подниматься над субстратом (поверхностный и воздушный мицелий). На воздушном мицелии обычно образуются органы размножения. Различают неклеточный (несептированный, или ценоцитический мицелий) лишенный перегородок и представляющий как бы одну гигантскую клетку с большим числом ядер, и клеточный (септированный) мицелий, разделенный перегородками – септами на отдельные клетки, содержащие от одного до многих ядер. Для представителей отделов хитридиомикотовых и зигомикотовых характерен неклеточный мицелий. У представителей отделов аскомикотовых и базидиомикотовых мицелий клеточный, с настоящими септами, образующими упорядочение, синхронно делению ядер. Септа развивается центрипетально, т. е. от стенки гифы к ее центру. В центре септы остается пора, через которую перемещаются питательные вещества, а также клеточные органеллы. У некоторых грибов, например дрожжей, вегетативное тело представлено одиночными почкующимися или делящимися клетками. Если такие почкующиеся клетки не расходятся, образуется псевдомицелий. Некоторые примитивные грибы имеют одноклеточный таллом, иногда лишенный клеточной стенки. Такие одноклеточные талломы грибов способны развивать ризомицелий – разветвленные нитевидные структуры, лишенные ядер. При формировании плодовых тел и некоторых вегетативных структур гифы грибов плотно переплетаются, образуя ложную ткань – плектенхиму. Основой вегетативного тела большинства грибов является мицелий, представляющий собой систему тонких ветвящихся нитей, или гиф, с апикальным ростом и боковым ветвлением. Различают мицелий не септированный (не клеточный), лишённый перегородок и представляющий собой как бы одну гигантскую клетку с огромным числом ядер, и септированный мицелий, разделённый септами (перегородками) на отдельные клетки. Отсутствие в мицелии септ является отличительным признаком низших грибов от высших. Многие грибы используются как продуценты антибиотических веществ. В последнее время установлено, что подавляющее количество видов грибов образуют биологически активные вещества, ферменты, органические кислоты, витамины, в связи с чем они являются перспективными объектами медицинской промышленности и фармакологии. Строение клетки. Клеточная стенка грибов содержит до 80–90 % полисахаридов, связанных с белками и липидами. Кроме того, в ее состав входят полифосфаты, пигменты, например меланины, и другие вещества. Микрофибриллярные скелетные компоненты кле- точной стенки состоят из хитина. Аморфный матрикс образован пре- имущественно глюканами с разными типами связей. Состав полисахаридов клеточной стенки специфичен для пред- ставителей крупных таксономических групп грибов. У хитридиоми- котовых, аскомикотовых и базидиомикотовых грибов в клеточных стенках содержатся хитин и глюканы. По этому признаку зигомико- товые резко отличаются от перечисленных отделов: клеточные стенки их мицелия содержат хитозан – полимер D-глюкозамина, но в них от- сутствуют глюканы. У грибов с дрожжеподобным талломом, вне за- висимости от их систематического положения количество хитина в клеточных стенках снижено, но содержатся маннаны – полимеры маннозы. В цитоплазме клеток настоящих грибов хорошо различимы рибо- сомы, митохондрии и типичные эукариотические ядра. Протопласт грибов окружен цитоплазматической мембраной – плазмалеммой. На границе между цитоплазмой и вакуолью также расположена мембра- на – тонопласт. Между тонопластом и плазмалеммой находится внутренняя мембранная система – эндоплазматическая сеть. Митохондрии настоящих грибов имеют пластинчатые кристы и сходны с митохондриями высших растений и животных. По современным представлениям типичный аппарат Гольджи от- сутствует, а его функцию выполняют различные видоизменения эндоплазматической сети. В растущих участках гиф в большом количе- стве содержатся пузырьки, или везикулы, происходящие от эндоплаз- матической сети. Они участвуют в транспорте веществ к месту синте- за клеточной стенки. В цитоплазме клеток грибов часто присутствуют микротельца – круглые или овальные мембранные структуры. Возможно, они иден- тичны или являются предшественниками лизосом или пероксисом – органелл, содержащих соответственно гидролитические ферменты или каталазу. В клетках грибов находится от одного до 20–30 ядер. Их размер обычно около 2–3 мкм. Они окружены оболочкой, состоящей из двух мембран. В нуклеоплазме содержатся ядрышко и хромосомы. При митотическом делении ядра ядерная оболочка часто сохраняется. В клетках грибов присутствуют многочисленные включения: гра- нулы гликогена, капли липидов. В вакуолях часто находятся гранулы белков и волютина. Подвижные клетки грибов – зооспоры и гаметы среди настоя- щих грибов характерны только для хитридиомикотовых. Жгутики на- стоящих грибов построены по типу, характерному для всех эукариот: их главный стержень (аксонема) состоит из двух одинарных цен- тральных микротрубочек, окруженных кольцом из девяти дублетов микротрубочек. Аксонемы у грибов соединяются с кинетосомой, или базальным телом, разными способами. У некоторых хитридиомико- товых передняя часть ядра и его бока окружены так называемым ядерным колпачком, а кинетосома почти полностью опоясана боль- шой полушаровидной митохондрией. Жгутики у настоящих грибов в отличие от псевдогрибов, бичевидные гладкие, расположены на зад- нем конце. На поверхности одиночных клеток некоторых грибов – спор го- ловневых и клеток дрожжей – обнаружены тонкие нитевидные струк- туры, названные фимбриями. Они имеют длину 0,5–10 мкм и тол- щину 6–7 нм. Предполагают, что фимбрии играют существенную роль в агрегации клеток. Грибы размножаются вегетативным, бесполым и половым путем Грибы размножаются вегетативным, бесполым и половым путем. При вегетативном размножении от мицелия отделяются неспециализированные его части, которые дают начало новому мицелию. Одной из форм вегетативного размножения можно считать и образование хламидоспор – толстостенных клеток, предназначенных для перенесения неблагоприятных условий. У дрожжей вегетативное размножение происходит путем почкования клеток. Бесполое размножение происходит при помощи специализированных неполовых клеток (реже многоклеточных структур) – спор. Споры у грибов развиваются эндогенно – внутри спорангиев или экзогенно – на особых веточках мицелия – конидиеносцах. У зигомицетов в спорангиях развиваются неподвижные споры, называемые спорангиоспорами. Спорангии находятся на специализированных гифах – спорангиеносцах, поднимающихся над субстратом. Конидии, как и спорангиоспоры, – неподвижные споры, пассивно переносимые токами воздуха, каплями дождя, насекомыми и другими способами. Однако в отличие от эндогенных спорангиоспор они образуются на мицелии или его специализированных ветвях – конидиеносцах экзогенно. Известно несколько типов формирования конидий из конидиогенных клеток. Конидиеносцы грибов имеют весьма разнообразное строение, часто наблюдается их агрегация в коремии, ложа или образование полузамкнутых структур – пикнид, в которых развиваются конидиеносцы и конидии. Бесполое размножение при помощи конидий характерно для базидиомикотовых и аскомикотовых, в том числе и анаморфных, где бесполое размножение достигло максимального разнообразия. Половое размножение отмечено у всех групп грибов, кроме анаморфных, называемых также несовершенными грибами. Формы полового процесса у грибов разнообразны. Их можно разделить на три большие группы: гаметогамия, гаметангиогамия и соматогамия.
Экологические группы грибов Почвенные грибы. Участвуют в минерализации органического вещества, образовании гумуса и т. п. В этой группе выделяют грибы, попадающие в почву только в определенные периоды жизни, и грибы ризосферы растений, живущие в зоне их корневой системы. Специализированные почвенные грибы: копрофиллы - обитающие на почвах, богатых перегноем (навозные кучи, места скопления помета животных); кератинофиллы - обитающи
|
|||||||||
|
|
Последнее изменение этой страницы: 2019-05-19; просмотров: 525; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 52.14.223.136 (0.015 с.) |






































 Существование планктонных организмов во взвешенном состоянии в воде обеспечивается некоторыми специальными приспособлениями. У одних видов образуются разного рода выросты и придатки тела — шипы, щетинки, роговидные отростки, перепонки. У других видов происходит накопление в теле веществ с удельным весом меньше единицы, например капель жира, газовых вакуолей и т. д. \
Существование планктонных организмов во взвешенном состоянии в воде обеспечивается некоторыми специальными приспособлениями. У одних видов образуются разного рода выросты и придатки тела — шипы, щетинки, роговидные отростки, перепонки. У других видов происходит накопление в теле веществ с удельным весом меньше единицы, например капель жира, газовых вакуолей и т. д. \



