Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Біологічна активність комплексів
Комплекс антиген-антитіло наділений вираженою біологічною активністю. Найбільш активні комплекси при помірному надлишку антигену. Вони споріднені з тканинами, взаємодіють з комплементом і можуть викликати в організмі наступні явища: дегрануляцію тучних клітин і базофілоцитів крові, підвищення вмісту ацидофілоцитів (еозинофілію), аглютинацію лейкоцитів і тромбоцитів, скорочення гладких м’язів, підвищення проникності капілярів, пошкодження нейтрофілоцитів, дегенерацію тканин, що адсорбують комплекс, а також лихоманку, інтоксикацію, феномен Артюса, шкірні реакції негайного і сповільненого типу. Зміни, які викликаються комплексом антиген-антитіло в тканинах, їх ступінь та динаміка розвитку залежать від кількості реагентів та особливостей антитіл. Вони можуть проявлятися у вигляді гострих запальних реакцій з некрозом в результаті адсорбції комплексом антиген-антитіло комплементу з наступним залученням нейтрофілоцитів, а також у вигляді хронічних запальних реакцій з гіаліновим потовщенням оболонки капілярів, внаслідок відкладення під нею комплексу антиген-антитіло без вираженої клітинної реакції. Системні пошкодження, які викликаються комплексом антиген-антитіло, частіше бувають сповільненого характеру і виражені слабо. Агреговані в результаті хімічних реакцій або нагрівання гамаглобуліни мають властивості, аналогічні властивостям комплексу антиген-антитіло. Нерозчинні комплекси антиген-антитіло викликають різноманітні алергічні реакції при надлишку антигену. При надлишку антитіл, коли в сироватці утворюється розчинний комплекс антиген-антитіло, реакції відсутні. Після повного видалення із сироватки комплексів антиген-антитіло у крові виявляються вільні антитіла. Питання для самоконтролю 1. Охарактеризуйте феномени та сили взаємодії реакції антиген-антитіло. 2. Опишіть фази реакції антиген-антитіло. 3. Поясніть, якою буде будова макроагрегата в залежності від співвідношення часточок антигена і антитіл. 4. Специфічність реакції антиген-антитіло. 5. Концентрація реагентів у реакції антиген-антитіло. 6. Особливості протікання реакції антиген-антитіло в залежності від характеристики реагентів (повні чи неповні антигени, повні чи неповні антитіла). 7. Якими методами можна розірвати комплекс антиген- антитіло? 8. Поясніть, які комплекси антиген-антитіло наділені найбільшою біологічною активністю.
Органи, тканини і клітини імунної системи
1. Загальна характеристика імунної системи Анатомічний синонім імунної системи – лімфатична система. Однак зрозуміти будову та функціонування імунної системи можна лише визначивши взаємозв’язки лімфатичної системи з іншими системами організму, принаймі із системою клітин крові та кровоносних судин, а також покривними тканинами (слизовими оболонками та шкірою). Ці системи – найближчі партнери, на які в своїй роботі опирається система лімфоцитарного імунітету. В організмі дорослої людини міститься близько 1013 лімфоцитів, тобто приблизно кожна 10 клітина тіла – лімфоцит. Як вони розміщені в організмі? Анатомо-фізіологічний принцип побудови імунної системи – органно-циркуляторний. Це означає, що є ряд спеціалізованих органів із організованою внутрішньою структурою (рис.12). При цьому лімфоцити не “сидять” в лімфатичних органах постійно (на відміну від, наприклад, гепатоцитів у печінці), а інтенсивно рециркулюють між лімфатичними органами та нелімфатичними тканинами через лімфатичні судини та кров. Міграція лімфоцитів з крові в тканини та з тканин до крові відбувається через стінку судин, і механізм цієї міграції включає в себе специфічну взаємодію певних молекул на мембрані лімфоцита з певними молекулами на мембрані клітин ендотелію стінки судин (такі молекули називають адгезинами, селектинами, інтегринами). Ці взаємодії відбуваються не в будь-якому місці судини, а в певних місцях, наприклад в лімфатичних вузлах – це ендотелій посткапілярних венул. Процес міграції лімфоцитів, звичайно, не носить характер випадкового переміщення, а строго регулюється рядом факторів, що залежать від місцевих тканинних та системних фізіологічних “задач” організму.
Рис. 12. Локалізація імунної (лімфоїдної) системи в організмі людини. 1 – кровотворний кістковий мозок; 2 – тимус; 3 - неінкапсульована лімфоїдна тканина слизових оболонок; 4 – лімфатичні вузли; 5 – судини лімфодренажу покривних тканин (аферентні лімфатичні судини); 6 – грудна лімфатична протока (впадає у системну циркуляцію (кров) через верхню порожнисту вену); 7 – селезінка; 8 – печінка; 9 – внутріепітеліальні лімфоцити.
Виділяють наступні органи та тканини імунної системи: 1. Кровотворний кістковий мозок – центральний орган всього кровотворення. Інкапсульовані органи 2. Тимус 3. Селезінка 4. Лімфатичні вузли Неінкапсульована лімфоїдна тканина слизових оболонок 5. Лімфатична тканина, асоційована зі шлунково-кишковим трактом. Це мигдалини, аденоїди, апендикс, пеєрові бляшки. Особливою субпопуляцією є внутріепітеліальні лімфоцити слизової оболонки кишки. 6. Лімфатична тканина, асоційована з бронхами, це субпопуляція слизової оболонки дихальної системи. 7. Лімфатична тканина інших слизових оболонок. 8. Особливі субпопуляції лімфоцитів у печінці, які в якості лімфоїдного бар’єру “обслуговують” кров ворітної вени. 9. Лімфоїдна підсистема шкіри, що включає в себе субпопуляцію особливих внутріепітеліальних лімфоцитів шкіри та регіонарні лімфатичні вузли та судини лімфодренажу. 10. Периферійна кров – транспортно-комунікаційний компонент імунної системи. Кровотворний кістковий мозок і тимус називають центральними органами імунної системи тому, що на їх території відбувається диференціація лімфоцитів із стовбурової кровотворної клітини, так званий лімфопоез. Лімфопоез – це диференціація лімфоцитів від стовбурової кровотворної клітини до зрілого неімунного лімфоцита. Зрілі неімунні лімфоцити локалізуються в периферійних лімфатичних органах та циркулюють між ними через кров. На території периферійних лімфатичних органів зрілі неімунні лімфоцити вступають в контакти з антигенпрезентуючими клітинами. Якщо антигенрозпізнаючий рецептор лімфоцита зв’язує комплементарний антиген на території периферійних лімфоїдних органів, де в нормі створюються усі необхідні умови для початку розвитку імунної відповіді, то лімфоцит вступає на шлях додиференціювання в режимі імунної відповіді, тобто починає проліферувати та продукувати ефекторні молекули (цитокіни, перфорин, цитолізини, гранзими та інші, в залежності від субпопуляції лімфоцита). Диференціювання лімфоцитів на периферії після розпізнавання антигена називають імуногенезом. Обов’язковим процесом на початку імуногенезу лімфоцитів в периферійних лімфатичних органах є проліферація клонів лімфоцитів, що розпізнали антиген. У результаті імуногенезу розвиваються клони імунних або ефекторних лімфоцитів, які в англомовній літературі називають armed (озброєними) або effector (ефекторними) лімфоцитами. Імунні лімфоцити розпізнають антиген та організовують його деструкцію в різних периферійних тканинах організму, де цей антиген присутній. У кістковому мозку відбувається диференціація усіх лейкоцитів крові. Після виходу із кісткового мозку в периферійні тканини лейкоцити в нормі вже ніколи не будуть проліферувати (на відміну від лімфоцитів). У відповідь на адекватні сигнали зовні вони лише будуть активовані до виконання своїх призначених в процесі диференціації функцій. На території кісткового мозку зі стовбурової кровотворної клітини утворюється загальна клітина – попередниця усіх лімфоцитів, з якої також на території кісткового мозку проходить поез 3 із 4 її нащадків: В-2 лімфоцити, нормальні кілери (NK) та дендритні клітини (DC). Четвертий нащадок, комітований (запрограмований) до диференціювання в Т-лімфоцити, мігрує для проходження поезу з кісткового мозку в тимус, і якась частина – в слизові оболонки, переважно кишково-шлункового тракту.
Які ж клітини входять до складу імунної системи? Істинні імуноцити – це всі варіанти лімфоцитів – Т, В, NK, DC. Безпосередні клітини-співробітники лімфоцитів – усі варіанти лейкоцитів – нейтрофіли, моноцити/макрофаги, еозинофіли, базофіли, тучні клітини. І навіть еритроцити вносять свій вклад у деструктивне завершення імунної відповіді – транспортують імунні комплекси антигена з антитілом і з комплементом (на еритроцитах є рецептори для комплементу) в печінку та селезінку для фагоцитозу та руйнування. Окрім клітин, “імунологічна матерія” представлена розчинними молекулами – гуморальними факторами. Це продукти В-лімфоцитів – антитіла (імуноглобуліни) і розчинні медіатори міжклітинних взаємодій – цитокіни. Цитокіни – це та молекулярна “матерія”, за допомогою якої лімфоцитарний імунітет “вбудований”, інтегрований в організм в цілому. Цитокіни – молекули, що секретуються клітинами у позаклітинне середовище з метою впливати на інші клітини або на себе ж, подавати сигнал до запуску тих чи інших процесів у клітинах-мішенях. 1. Органи імунної сиситеми 1.1. Тимус. Тимус – спеціалізований лімфоїдний орган, в якому проходить лімфопоез більшість Т-лімфоцитів організму. Тимус розміщений в передньому верхньому середньостінні, за грудиною, над серцем. Тимус складається з двох великих доль, які фрагментовані на велику кількість дольок, розділених фіброзними перегородками (рис. 13). Ці дольки і є структурними одиницями будови тимусу. У кожній дольці чітко помітні дві гістологічні зони: по периферії – коркова, в центрі – медулярна. Строма тимусу – епітеліальна.
Рис. 13. Схема будови дольки тимусу. 1 – коркова зона; 2 – мозкова речовина; 3 – епітеліальні клітини кори (nurse cells) (ектодермального походження); 4 – епітеліальні клітини мозкової речовини (ентодермального походження); 5 – дендритні клітини (кістковомозкового походження); 6 – макрофаги (кістковомозкового походження); 7 – тимоцити – лімфоцити тимусу (кістковомозкового походження).
Епітеліальні клітини тимусу мають особливу морфологію. Епітеліальні клітини кори своїми відростками “обнімають та колихають” лімфоцити тимусу (тимоцити), тому вони називаються nurse cells (медсестри, нянечки). Епітеліальні клітини тимусу продукують цитокіни. Також на клітинах епітелію тимусу експресовані молекули адгезії, які комплементарні молекулам адгезії на тимоцитах. Ці взаємодії і втримують тимоцити, що розвиваються на території тимусу на необхідний для диференціювання час. Клітини мезодермального або кістковомозкового походження в тимусі представлені тимоцитами, а також дендритними клітинами тимусу і макрофагами. Дендритні клітини розміщені переважно в зоні, перехідній між корковою і медулярною. Макрофаги присутні в корковій зоні, пограничній і медулярній. Тимоцити диференціюються із загальної стовбурової кровотворної клітини. Клітини-попередниці приходять в тимус через стінку великих венул в кортико-медулярній області і звідти мігрують у субкапсулярну зону. У мозковій зоні дольок є щільні утворення зі скручених епітеліальних клітин – тільця Гессаля (тільця вилочкової залози). Вірогідно, це місця компактного накопичення дегенеруючих клітин. Від інших лімфоїдних органів тимус відрізняє особлива постнатальна динаміка його морфогенезу в залежності від віку. До моменту народження тимус повністю сформований. Він густо заселений лімфоцитами (тимоцитами) протягом всього дитинства і до моменту статевого дозрівання. Після пубертатного періоду тимус починає зменшуватися в розмірах, зморщуватися. Тимусні гормони. Дозрівання лімфоцитів, які прибувають із кісткового мозку до тимусу, регулюється різними пепидами, що вважаються тимусними гормонами. Під впливом цих гормонів, незрілі лімфоцити утворюють поверхневі приймачі і позначки, дозрівають і диференціюються в Т-лімфоцити. Витяжки з тимусової залози, з Т-лімфоцитів і навіть з сироватки крові показали наявність тимусних гормонів; деякі з них були очищені до рівня, який дозволяє розпізнати їх властивості і дію. Гуморальний фактор тимусу був знайдений у телячому тимусі та ідентифікований як топоселективний протеїн, що складається з 31 амінокислоти. Цей фактор сприяє розмноженню Т-клітин, стимулює реакції Т-лімфоцитів до лектинів. Тимузин, другий гормон тимусу, в дійсності є сумішшю, що складається із 15 або більше протеїнів (гормонів); найвідомішим з них є тимузин-альфа1, теплостабільний пептид. Він посилює реакції Т-лімфоцитів до пектинів, спричиняє дозрівання Т-лімфоцитів. Тимузин-бета4 складається з 43 амінокислот. Тимулін – найменший з тимусних гормонів, складається лише з 9 амінокислот. Тимулін спричиняє появу Thy 1 антигена. Тимопоетин ІІ – найбільший із тимусних гормонів; він складається із 49 амінокислот. Цей гормон провокує продукцію Thy 1 антигена на недозрілих Т-лімфоцитах. 1.2. Фабрицієва сумка. Фабрицієва сумка – це лімфатична залоза, яка знаходиться на термінальному кінці кишок у птахів. Фабрицієва сумка – подовгастий орган, який у домашньої птиці атрофується після 4 місяця життя. Як і тимус, Фабрицієва сумка поділена на фолікули, у яких є лімфоцити, макрофаги і плазматичні клітини (рис. 14). Клітини Фабрицієвої сумки відрізняються від тимусних лімфоцитів поверхневим антигеном; якщо лімфоцити тимусу носять Thy 1 антиген, лімфоцити Фабрицієвої сумки носять В-антиген, який вони отримують у Фабрицієвій сумці. Лімфоцити птахів утворюються в тимусі і Фабрицієвій сумці; назва В-антигена походить від слова “бурза”, яка є місцем дозрівання лімфоцитів.
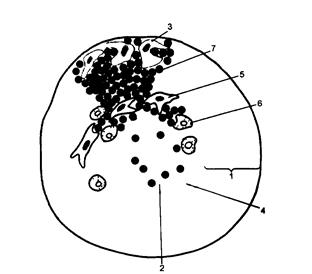
Рис. 14. Анатомічне взаємовідношення Фабрицієвої сумки та клоаки у курчати. 1 – протока Фабрицієвої сумки; 2 – анус; 3 – клоака; 4 – Фабрицієва сумка.
Ссавці не мають Фабрицієвої сумки. Але мають В-лімфоцити, які дозрівають у бурзо-еквівалентних тканинах. Такі лімфатичні органи асоціюються із кишковим трактом і кістковим мозком; прикладом такої асоціації є лімфатичні острівці Пеєра. Хірургічне видалення бурзи у птахів чи кишковоспоріднених лімфатичних тканин у ссавців зменшує кількість В-лімфоцитів і пригнічує продукцію антитіл. Імунодефіцит можна створити тоді, коли хірургічні видалення лімфатичних тканин проводяться у молодих тварин, в яких периферійні лімфатичні органи потрібні не тільки для надання лімфоцитам спеціальних антигенів, але також вони є місцем розмноження лімфоцитів та їх імунного дозрівання. 1.3. Лімфатичні вузли. Лімфатичні вузли – чисельні, симетрично розміщені по тілу, інкапсульовані периферійні лімфоїдні органи бобовидної форми, розміром від 0,5 до 1,5 см в довжину (не запалені). Лімфатичні вузли через аферентні лімфатичні судини (яких декілька на один вузол) дренують тканинну рідину з усіх бар’єрних тканин. Лімфатичні вузли розміщені регіонарно і називаються у відповідності з частиною тіла, яку вони “обслуговують”: навколовушні, задньошийні, підпахвинні, підколінні і т.д. Таким чином, лімфатичні вузли – це “таможня” для всіх речовин (антигенів), що потрапляють у субкапсулярний синус лімфатичного вузла. Із анатомічних воріт вузла паралельно з артерією і веною виходить єдина еферентна судина, що несе лімфу в грудну лімфатичну протоку, яка впадає в нижню порожнисту вену і таким чином лімфа вливається в системний кровотік. Лімфатичний вузол має коркову та медулярну зони (рис. 15). Коркова зона розділена трабекулами на радіальні сектори. У цій зоні розміщуються лімфоїдні фолікули – В-лімфоцитарна зона. Лімфатичні фолікули проходять 3 стадії розвитку, які називають по-різному. Первинний фолікул – дрібний фолікул, що складається з неімунних В-лімфоцитів. Після того, як В-лімфоцит розпізнає антиген, отримає усі необхідні стимулюючі сигнали, він вступить в імуногенез, строго необхідним етапом якого є проліферація клону В-лімфоцитів. Фолікул, що містить інтенсивно проліферуючі В-лімфоцити, називається гермінативним центром. Первинний фолікул перетворюється в гермінативний центр протягом приблизно 1 тижня після активної імунізації.
Рис. 16. Схема будови лімфатичного вузла. 1 – аферентні лімфатичні судини; 2 – еферентна лімфатична судина; 3 – артерія; 4 – вена; 5 – посткапілярна венула у паракортикальній зоні; 6 – первинний фолікул; 7 – гермінативний центр; 8 – вторинний фолікул (фолікули первинні, гермінативні центри і вторинні – це В-клітинна зона); 9 – паракортикальна зона – Т-клітинна (тут локалізовані посткапілярні венули з високим ендотелієм); 10 – мозкова речовина.
Після завершення процесу імуногенезу фолікул суттєво зменшується в розмірах, в цей період його називають вторинним фолікулом. У паракортикальній зоні лімфатичного вузла локалізовані Т-лімфоцити і посткапілярні венули, через стінку яких відбувається міграція лімфоцитів з крові у лімфатичний вузол. Це Т-залежна зона. В активному стані посткапілярні венули мають особливий за морфологією ендотелій – високий. У Т-залежній зоні міститься багато інтердигітальних дендритних клітин. Інтердигітальні дендритні клітини – це клітини кістковомозкового походження, які мігрували у вузол з тканинною рідиною по аферентних лімфатичних судинах з покривних тканин (у шкірі ці тканини називають клітинами Лангерса), вони є антигенпрезентуючими для Т-лімфоцитів. Під паракортикальною зоною розміщені медулярні тяжі, у яких багато макрофагів, і, якщо вузол знаходиться у стані активної імунної відповіді, в медулярних тяжах можна бачити плазматичні клітини. Медулярні тяжі впадають в медулярний синус, з якого виходить еферентна лімфатична судина. 1.4. Печінка. Печінка має особливі функції в імунітеті. Саме в печінці, в синусоїдах у людини локалізована більша частина особливих лімфоїдних клітин – нормальних кілерів (NK). Крім того, в печінці є особливі субпопуляції Т-лімфоцитів. Є обґрунтоване припущення, що лімфоцити печінки забезпечують постійне підтримання імунологічної толерантності до харчових речовин і імунна система не витрачає себе таким чином на щоденні імунні відповіді на харчові речовини. І, нарешті, в печінці знаходиться мало не половина маси усіх тканинних макрофагів організму. У синусоїдах печінки, так само як і в синусоїдах селезінки, макрофаги фагоцитують і розщеплюють імунні комплекси (комплекси антигенів з антитілами і білками комплементу), які приносять сюди на собі старіючі еритроцити. 1.5. Селезінка. Подібно до того, як лімфатичні вузли фільтрують і очищують лімфу, так селезінка очищує кров. Селезінка знаходиться в черевній порожнині і має видовжену форму. Важливість селезінки полягає в тому, що вона забирає із крові використані клітини (зокрема еритроцити), антигени чи мікроби і її клітини продукують імуноглобуліни. У осіб похилого віку селезінка не так активно проявляє свої імунні функції. Раніше, коли не було можливості пригнічувати імунну реактивність організму медикаментозно, практикували хірургічне видалення селезінки. Таке видалення дещо зменшувало імунні реакції і допомагало у пригніченні аутоімунності. Селезінка поділена на дві зони. Одна зона називається червоною пульпою, і в ній акумульовані еритроцити; друга - білою пульпою, і у ній наявні лімфоцити та відбуваються імунні реакції. Вздовж артерій селезінки є лімфатичні тканини, які мають Т- і В-лімфоцити. При антигенній стимуляції ці клітини розмножуються і утворюють проростаючі центри. Біла пульпа відділена від червоної зоною, складеною переважно із макрофагів. Велика частина внутрівенно введеного антигена затримується у селезінці; її макрофаги фагоцитують чи піноцитують антигени і перетворюють їх у імуностимулюючі антигени; у білій пульпі трансформовані антигени стимулюють відповідні В-лімфоцити, які утворюють проростаючі центри і продукують антитіла. Деякі лімфоцити трансформуються в пам'ятаючі клітини і переносяться до кісткового мозку. Тому при другій стимуляції тим самим антигеном багато антитіл класу IgG утворюються в кістковому мозку, бо там є пам'ятаючі клітини. 1.6. Другорядні лімфатичні органи. У дітей часто піддаються інфекції лімфатичні тканини, наявні в мигдалевидних залозах. Ці залози схожі до тимусу, тому що активні у дітей, а в дорослому віці значно зменшуються і частково атрофуються. Відрізняються ці залози від тимусу наявністю В, а не Т-лімфоцитів. Хірургічне видалення цих залоз хоч не приносить великої шкоди, не корисне для імунного стану людини. Подібним другорядним лімфатичним органом є апендикс, який має В- і Т-лімфоцити.
2. Лімфоїдні клітини 2.1. Лімфоцити. В імунологічній відповіді беруть участь В-лімфоцити, Т-лімфоцити і макрофаги. Усі клітини імунної системи, що відповідають за імунну реактивність і неспецифічну резистентність, виникають з єдиної гемопоетичної стовбурової клітини. Диференціація не має зворотного ходу, тобто, якщо клітина пройшла певний етап диференціації, вона вже не здатна диференціюватися в іншому напрямку або повернутися на попередній етап. Лімфоцити беруть участь у всіх імунологічних реакціях і є центральною ланкою імуногенезу. Вони здатні реагувати лише на окрему групу структурно подібних антигенів. Ця здатність існує ще до першого контакту імунної системи з даним антигеном і зумовлена наявністю мембранних рецепторів до його детермінант. Кожен клон лімфоцитів відрізняється від іншого будовою цих рецепторів. Таким чином, кожен клон здатний реагувати тільки на певні, відповідні йому антигени. В результаті лімфоцити людини складають виключно неоднорідну популяцію клітин, у яких різноманіття рецепторів перевищує 106. Лімфоцити відрізняються між собою не тільки за специфічністю своїх рецепторів, а й за функціональними властивостями. Розрізняють два основних класи лімфоцитів: В-лімфоцити й Т-лімфоцити. Лімфоцити сприймають антигенні подразнення, здатні диференціюватися в клітини-ефектори, проліферувати, збільшуючи масу клітин, що реагують на даний антиген, зберігають імунологічну пам’ять, забезпечують виникнення імунологічної толерантності, виділяють медіатори. Основними властивостями лімфоцитів є: рухливість, висока чутливість до дії фізичних, хімічних та біологічних факторів і здатність диференціюватися під впливом специфічного антигена. Вони здатні проникати в цитоплазму деяких типів епітеліальних клітин. Лімфоцити з крові виходять в лімфатичні вузли через кубовидний епітелій посткапілярних венул. Вважається, що кубовидний епітелій здатний притягувати лімфоцити. Лімфоцити складають основну масу клітинних елементів лімфоїдної тканини, 95% клітин лімфи та 30% загальної кількості лейкоцитів крові. Лімфоцити складають значну частку клітин кісткового мозку. З кісткового мозку частина їх поступає в кровотік, проникаючи в судини між епітеліальними клітинами синусів. Вони мігрують у тимус, селезінку, лімфатичні вузли, а також в мієлоїдні центри. Певна кількість стовбурових клітин кісткового мозку постійно мігрує в кров. Стовбурові клітини, що поступають у тимус, набувають імунологічної компетентності та забезпечують постійну можливість відповіді на нові антигенні стимули. Якщо опроміненим тваринам ввести внутрівенно клітини кісткового мозку, що містять хромосомну мітку, то першочергово вони виявляються тільки в тимусі, а в потім – в селезінці та лімфатичних вузлах. Лімфоцити поступають в кров через грудну протоку. Через ендотелій посткапілярних венул лімфовузлів та селезінки вони мігрують в тканину цих органів, пересікають лімфоцитарні поля і по еферентних шляхах залишають їх. Далі вони знову поступають в кров через грудну протоку. У людини тривалість життя циркулюючих лімфоцитів вираховується місяцями чи навіть роками. Протягом всього життя довгоживучі лімфоцити сотні разів циркулюють з лімфовузлів та інших лімфоїдних органів у лімфу та кров і знову в органи. Поява у філо- та онтогенезі у хребетних виражених специфічних імунних реакцій співпадає з появою лімфоцитів та лімфоїдної тканини. Зменшення кількості лімфоцитів внаслідок тимектомії, впливу іонізуючих випромінювань або дренування протягом декількох днів грудної протоки приводить до послаблення специфічної імунологічної реактивності як гуморального, так і клітинного типу. Імунологічна реактивність у цих випадках може бути відновлена введенням сингенних лімфоцитів. Якщо вводити лімфоцити тварин, толерантних до певного антигена, то реактивність опромінених тварин відновлюється по відношенню до гетерогенних антигенів, але відсутня до гомологічного. Ці дані показують, що лімфоцити є також клітинами, відповідальними за стан імунологічної толерантності. 2.2. Плазмоцити. Утворення антитіл плазмоцитами сьогодні вже є твердо доведеним фактом. Синтез антитіл плазмоцитами здійснюється, в основному, в лімфатичних вузлах, селезінці та кістковому мозку. Плазмоцит проходить наступні етапи розвитку: плазмобласт (15-20 мкм), юний плазмоцит (10-15 мкм) і зрілий плазмоцит (10-12 мкм). З регіонарних лімфатичних вузлів плазмобласти мігрують в інші лімфатичні органи, де диференціюються в плазмоцити, що забезпечує генералізацію імунної відповіді. Плазмоцити резистентні до іонізуючих променів. Так, опромінення плазмоцитів дозами до 100 Гр не впливає на синтез антитіл. Тривалість життя більшості плазмоцитів в межах 48 годин, однак частина їх зберігається в організмі до 6 місяців. У зрілому плазмоциті виявляється 1,25 х 1013 – 7,5 х 1013 антитіл. За секунду клітина може продукувати 1000 – 2000 молекул антитіл (приблизно сто мільйонів молекул антитіл за добу). Плазмоцити синтезують антитіла лише до однієї антигенної детермінанти одного класу та субкласу і лише одного алотипічного варіанту. Рецептори клітин-попередниць за своєю специфічністю та приналежністю до класу, підкласу, алотипу та ідіотипу аналогічні тим антитілам, які буде виробляти дана клітина після антигенної стимуляції. Звільнені від рецепторів лімфоцити називають “голими”. При інкубуванні з імуноглобулінами іншої специфічності лімфоцити адсорбують їх на своїй поверхні і вони стають рецепторами. Такі нові рецептори сприймають антигенний стимул та індукують синтез антитіл, але специфічних не до другого рецептора, а до першого, тобто до того, до якого клітина предетермінована. Якщо видалити рецептори з поверхні клітин, предетермінованих до синтезу людського гамаглобуліну, і “голі” лімфоцити інкубувати з рецепторами для бичачого сироваткового альбуміну, то при стимуляції цих клітин бичачим сироватковим альбуміном клітини синтезують антитіла до людського гамаглобуліну. Плазмоцити у лімфоїдній тканині утворюють окремі клони, що виникли в результаті проліферації та диференціювання родоначальної клітини. Характерна особливість клону – здатність продукувати антитіла до певного антигена. Ця здатність набута родоначальними клітинами клону, індукованими даним антигеном, і передається усім клітинам клону. В клоні одночасно виявляються плазмоцити різного ступеню зрілості. Це говорить про те, що інформація, яка зумовлює їх здатність виробляти специфічні антитіла, передається генетично від плазмобласта зрілому плазмоциту. Підтвердженням такого припущення є першочергова поява в процесі імуногенезу антитілотвірних плазмобластів, а потім наступне збільшення кількості уже зрілих форм плазмоцитарного ряду, а також залежність імунної відповіді на даний антиген не від кількості зрілих плазмоцитів, а їх попередників. Завдяки механізмам, що забезпечують можливість постійної зміни клонів імунокомпетентних клітин, чутливих до того чи іншого антигена, зберігається постійна готовність лімфоїдної тканини реагувати на новий антиген, що поступає в організм. При первинній відповіді утворюється відносно невелика кількість плазмоцитів, але кількість їх різко збільшується при повторних імунізаціях. При первинній стимуляції кроликів антигеном сальмонел плазмоцити виявляються в регіонарному лімфовузлі на 4 добу, кількість їх подвоюється на 6-8 добу, а на 14 добу падає до вихідного рівня. При вторинній відповіді вони з’являються вже через 2 доби, а в період між 2-4 добами кількість їх подвоюється кожних 12 годин. При первинній відповіді плазмоцити виробляють IgM, а при вторинній – IgG. Клітини, що виробляють IgM i IgG, не відрізняються між собою морфологічно. Існують клітини - лімфоплазмоцити, що вже не виробляють антитіла, але здатні під впливом антигенного стимулу знову почати їх синтез. Ці клітини виявляються переважно в синусних лімфовузлах. 2.3. В-лімфоцити. Попередники антитілопродукуючих клітин розвиваються із гемопоетичної стовбурової клітини кісткового мозку. При цьому вони проходять свій цикл розвитку, в результаті якого на їх поверхні з’являються рецептори, спочатку у вигляді молекул IgM, а в міру дозрівання виникають IgD, рецептори компонентів комплементу, Fc-фрагментів тих імуноглобулінів, які вони здатні продукувати. Всередині кожного клону частина В-клітин переключається із синтезу IgM (IgD) на синтез IgG, IgA, IgE. Зрілі В-клітини відзначаються тим, що на їх мембранах є імуноглобулінові рецептори до антигена. При зв’язуванні антигена цими рецепторами, клітини активуються, проліферують і диференціюють у плазмоцити, які є продуцентами антитіл. Основна функція В-лімфоцитів – участь у гуморальній імунній відповіді. Містяться у крайовій зоні білої пульпи селезінки та у зовнішній зоні кортикального шару лімфовузлів, де формуються зародкові центри фолікулів. Серед В-лімфоцитів можна вирізнити ряд субкласів. Основним субкласом є В-лімфоцити – попередники антитілотвірних клітин (плазмоцитів). Внаслідок їхньої активації антигеном та рядом специфічних та неспецифічних сигналів відбувається проліферація і диференціація клонів плазматичних клітин, які синтезують антитіла – імуноглобуліни. В-супресори пригнічують проліферацію В- і Т-лімфоцитів, разом з Т-супресорами відповідають за розвиток імунологічної толерантності. В-кілери можуть взаємодіяти з Fc-фрагментами антитіл, фіксованих на клітинах, які спричиняють руйнування цих клітин. В-клітини пам’яті формуються з частини стимульованих антигеном В-лімфоцитів. Ці клітини не диференціюються до кінця, а переходять у стан спокою на тривалий час. При повторному контакті з антигеном вони швидко перетворюються на антитілотвірні клітини і забезпечують імунологічну пам’ять. 2.4. Т-лімфоцити та їх різновиди. Попередники Т-лімфоцитів із кісткового мозку й ембріональної печінки мігрують у тимус і зазнають серії перетворень. Т-лімфоцити у фізіологічних умовах містяться навколо артеріол у білій пульпі селезінки, у паракортикальній зоні лімфовузлів. Основна функція Т-лімфоцитів – розпізнання антигену, спочатку переробленого і представленого на поверхні антигенпрезентуючих клітин. Т-лімфоцити відповідають за формування клітинного імунітету, а також допомагають В-лімфоцитам при гуморальній імунній відповіді. В процесі дозрівання і проліферації виникають різні види Т-лімфоцитів. Залежно від рівня і напрямку диференціювання Т-лімфоцити набувають певного набору мембранних маркерів, які визначаються моноклональними антитілами. Ці маркери є глікопротеїдами, їх позначають CD (кластер диференціювання). Зрілі Т-лімфоцити можуть мати CD4 або CD8, а також CD3. Т-лімфоцити розпізнають антиген і реагують на нього за допомогою Т-клітинного рецептора разом з молекулою CD3. Глікопротеїдний Т-рецептор міститься на мембрані Т-лімфоцита, складається з двох (альфа і бета) ланцюгів і реагує специфічно з епітопом антигену. Основні субпопуляції Т-лімфоцитів. Розрізняють Т-лімфоцити-хелпери (помічники) з CD4 антигеном, Т-супресори, Т-ефектори (кілери) з CD8 антигеном, Т-лімфоцити контрсупресори. Серед Т-хелперів теж є різновиди: класичні Т-хелпери, які активуються, проліферують під впливом антигена і стимулюють В-лімфоцити до поділу і перетворення їх у плазмоцити; Т-хелпери гіперчутливості сповільненого типу, що здійснюють функцію помічників шляхом вивільнення біологічно активних речовин – лімфокінів; Т-хелпери-підсилювачі – стимулюють інші Т-лімфоцити до продукції факторів, які виконують роль помічників. Таким чином, основною функцією Т-хелперів є стимуляція В-лімфоцитів і макрофагів. До клітин, що здатні пригнічувати різні ланки імунопоезу, відносяться Т-супресори (також мають маркер CD8+). Вони регулюють інтенсивність імунної відповіді, пригнічуючи активність CD4+-клітин, запобігають розвитку аутоімунних реакцій, забезпечують природну імунологічну толерантність до власних антигенів і організму матері до батьківських антигенів плода.Від функціонального стану Т-супресорів залежить розвиток алергічних, імунодефіцитних станів, вираженість трансплантаційних реакцій тощо. Розрізняють специфічні та неспецифічні Т-супресори. Специфічні Т-супресори пригнічують імунну відповідь організму на конкретні антигени, в той час, як неспецифічні супресори пригнічують відповідь незалежно від виду антигена. Наприклад, при функціональній недостатності Т-супресорів В-лімфоцити й Т-кілери одержують широку можливість реагувати проти власних клітин і тканин, що зумовлює розвиток аутоімунних і алергічних реакцій. Однак, якщо активність Т-супресорів переважає функціональну здатність Т-хелперів, створюється благодатний фон для розвитку імунодефіцитних станів. Особливу роль відіграють Т-контрсупресори (не мають ні CD8+, ні CD4+) Вони здатні пригнічувати вплив Т-супресорів на Т-хелпери. Важливою групою Т-лімфоцитів є Т-ефектори, кілери (цитотоксичні Т-лімфоцити). Вони мають мембранні маркери CD8+ і здатні спричинювати лізис клітин, які несуть на поверхні чужорідні антигени (клітини, інфіковані вірусом, або які мають мікробні антигени, клітини алотрансплантату тощо) - знищувати ці клітини разом з патогенними агентами, що в них знаходяться. Т-лімфоцити-зберігачі імунологічної пам’яті формуються під час первинної імунної відповіді і мають маркер CD4+. Розрізняють малоіснуючі (місяці) і довгоіснуючі (роки) клітини пам’яті. 2.5. Натуральні кілери. У крові людини на долю Т-лімфоцитів припадає біля 75% лімфоцитів, 15% складають В-лімфоцити і 10% - клітини, що не мають ознак ні Т-, ні В-клітин. Це так звана третя популяція клітин (нульові лімфоцити). Серед них вирізняють К-клітини (кілери), які здійснюють антитілозалежну цитотоксичність, тобто їхня кілерна дія реалізується внаслідок активації комплексом антиген-антитіло. Вони знищують опсонізовані антитілами (головним чином класу IgG та IgМ) клітини, переважно бактеріальні. Але є й нульові лімфоцити, які не активуються комплексом антиген-антитіло. Це природні, або натуральні кілери (НК або NK). Деякі дослідники вважають, що природні кілери є головним неспецифічним фактором протипухлинного захисту. Вони мають неспецифічну протипухлинну активність, розпізнають будь-яку пухлинну клітину та знищують її внаслідок з’єднання з її поверхнею та продукції перфоринів. Перфорини вкорінюються в мембрану пухлинної клітини та призводять до появи в клітинній оболонці „діри”, через яку відбувається прямий обмін між цитоплазмою та зовнішнім середовищем. Клітина гине. Натуральні кілери мають саме неспецифічну протипухлинну активність, вони розпізнають будь-яку пухлинну клітину без попередньої імунної відповіді проти її антигенів.
|
|||||||||
|
|
Последнее изменение этой страницы: 2017-02-10; просмотров: 493; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.111.183 (0.054 с.) |