
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Кафедра водных биоресурсов и аквакультурыСодержание книги Поиск на нашем сайте
Кафедра водных биоресурсов и аквакультуры
М е т о д и ч е с к и е у к а з а н и я по дисциплине «ИХТИОЛОГИЯ» (для студентов зооинженерного факультета, Водные биоресурсы и аквакультура»
САНКТ-ПЕТЕРБУРГ 2 0 1 1
УДК: 577.4:591.524.12 Методические указания рассмотрены на заседании методической комиссии зооинженерного факультета (протокол № от 2011года) и рекомендованы к печати
Составители: Н.Б. Рыбалова –канд. с-х. наук, доцент кафедры «Водные биоресурсы и аквакультура»; Г. Талалай –канд. биол. наук, доцент кафедры «Водные биоресурсы и аквакультура» СПб ГАУ; .
. Рецензенты: ОГЛАВЛЕНИЕ ВВЕДЕНИЕ ЧАСТЬ I. ОБЩАЯ ИХТИОЛОГИЯ 1.1. Место рыб в системе животных 1.2. Строение и физиологические особенности рыб 1.2.1. Форма тела и способы движения 1.2.2. Кожа, чешуя и органы свечения 1.2.3. Скелет рыб 1.2.4. Мышечная система и электрические органы 1.2.5. Пищеварительная система рыб 1.2.6. Дыхательная система и газообмен 1.2.7. Кровеносная система рыб 1.2.8. Выделительная система рыб и осморегуляция 1.2.9. Половая система рыб 1.2.10. Нервная система рыб и органы чувств 1.2.11. Железы внутренней секреции 1.2.12. Ядоносность и ядовитость рыб
ВВЕДЕНИЕ Ихтиология – наука о рыбах (от греч. «ихтиос» - рыбы, «логос» - слово, учение). Наиболее древние из известных суждений о рыбах как своеобразной группе живых организмов известны из письменных источников Индии (6 в. до н. э.), Китая (5 в. до н. э.), Древней Греции (4 в. до н. э.). Аристотель впервые выделил рыб в самостоятельную группу водных животных. В самостоятельный раздел научных исследований ихтиология по-настоящему начала оформляться только в 19 веке. В России наибольшее значение в этом плане имели научно-промысловые изыскания К. М. Бэра, Н. Я. Данилевского, Н. М. Книповича и других учёных на Каспийском, Азовском, Чёрном, северных и дальневосточных морях и в их бассейнах. Огромное значение имели труды К. Ф. Кесслера (промысловая ихтиология), Л. С. Берга (систематика, распространение), А. Н. Северцова (анатомия), В. В. Васнецова (морфология, эмбриология), Е. К. Суварова (физиология), Г. В. Никольского (частная ихтиология, экология, динамика численности рыб) и многих других русских и советских ихтиологов.
Из ихтиологии выделились такие науки и дисциплины, как физиология рыб, рыбоводство, промышленное рыболовство, эмбриология рыб, сырьевая база рыбной промышленности, технология рыбных продуктов, болезни рыб. Ихтиология изучает внешние признаки и внутреннее строение рыб (морфологию и анатомию), отношение рыб к внешней среде – неорганической и органической (экологию, иногда называемую биологией), историю развития – индивидуальную (эмбриологию) и историю развития видов, родов, семейств, отрядов, и так далее (эволюцию или филогению), наконец географическое распространение рыб (зоогеографию). Кроме того, ихтиология изучает закономерности колебания и численности стад рыб, разрабатывает способы определения их промысловых запасов, даёт краткосрочные и долгосрочные прогнозы уловов. Ихтиология изучает этологию рыб, их ориентацию, средства общения, формы заботы о потомстве. Разведение рыбы в сложных условиях комплексного использования водных ресурсов требует отглубокого знания основ ихтиологии: биологии рыб, строения их организма, видовых особенностей, реакций организма на изменение условий внешней среды. Методические указания по дисциплине «Ихтиология» предназначены для студентов зооинженерного факультета, специализирующихся по рыбоводству, специальность зоотехния. Они составлены в соответствии с программой курса «Ихтиология».
ЧАСТЬ I. ОБЩАЯ ИХТИОЛОГИЯ Строение и физиологические особенности рыб Строение и функции организма рыбы отражают его связь с водной средой. Специфические приспособления рыб весьма многообразны. Возникновение и характер их обусловлены чрезвычайным разнообразием среды и образа жизни. Скелет рыб Цель занятия: изучить скелет рыб. Материалы: плакаты, таблицы. На рис. 10. представлена схема скелета большей части рыб.
Рис. 10. Схема скелета рыб
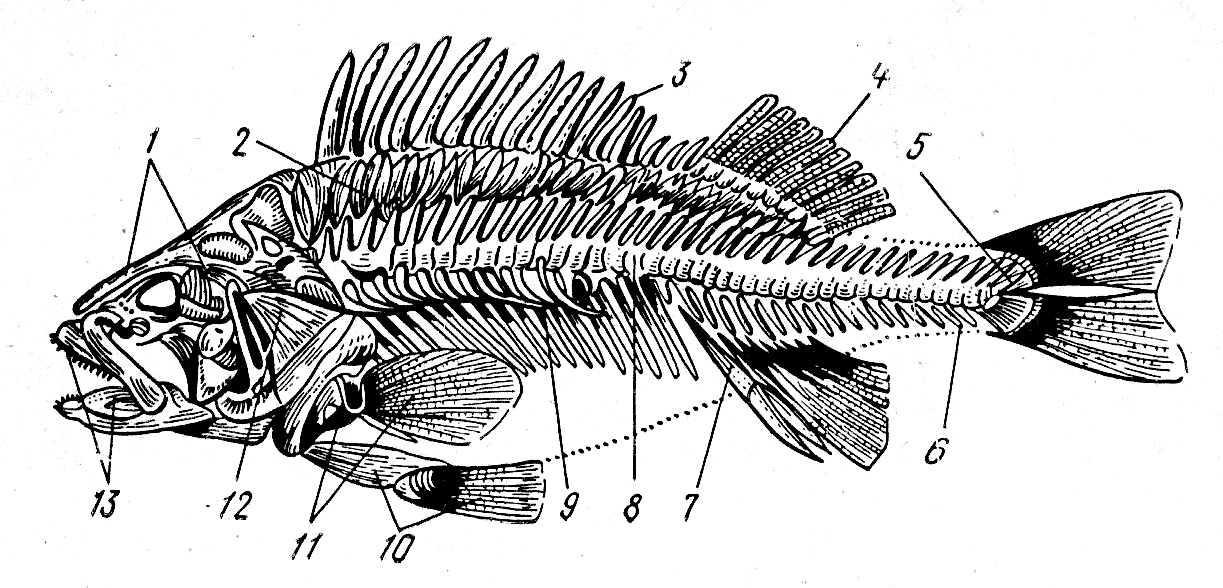
На рис. 11 представлен скелет костистой рыбы.
Рис. 11. Скелет костистой рыбы (окуня):
1 – кости черепа; 2 – основные элементы спинного плавника; 3, 4 – лучи спинного плавника; 5 – последние позвонки, удерживающие хвостовой плавник; 6 – хвостовые позвонки; 7 – основные элементы анального плавника; 8 – туловищные позвонки; 9 – рёбра с придатками; 10 – кости и лучи брюшного плавника; 11 – кости и лучи грудного плавника; 12 – жаберная крышка; 13 – верхняя и нижняя челюсти Внутренний скелет Осевой скелет может быть представлен хордой или позвоночником (табл. 11). Таблица 11 Виды осевого скелета рыб
Скелет головы представлен черепной коробкой (предохраняющей головной мозг и связанной с жаберным и челюстным аппаратом) табл. 12. Таблица 12 Скелет головы
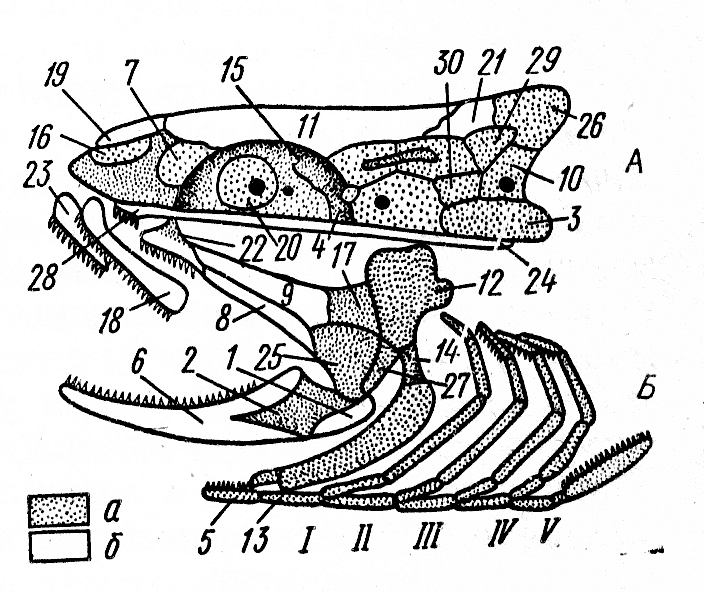
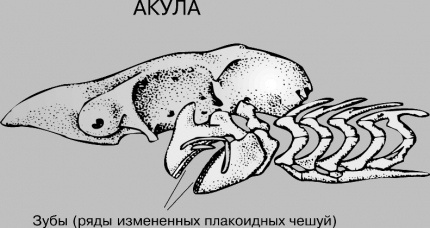
На рис. 12 ознакомтесь с представленными костями черепа костистой рыбы, на рис. 13 представлен череп и зубы судака и на рис. 14 череп и зубы акулы.
Рис. 12. Расположение костей в черепе костистой рыбы: А – висцеральный скелет; Б – мозговой (жаберная крышка не нарисована): а – основные кости и хрящ; б – покровные кости; 1 – угловая; 2 – сочленовная; 3 – основная затылочная; 4 – основная клиновидная; 5 – копула; 6 – зубная; 7 – боковая обонятельная; 8 - наружная крыловидная; 9 – внутренняя крыловидная; 10 – боковая затылочная; 11 – лобная; 12 – подвесок; 13 – гиоид; 14 – окостеневшая связка; 15 – боковая клиновидная; 16 - средняя обонятельная; 17 – задняя крыловидная; 18 – верхнечелюстная; 19 – носовая; 20 - глазоклиновидная; 21 – теменная; 22 – нёбная; 23 – предчелюстная; 24 – парасфеноид; 25 – квадратная; 26 – верхняя затылочная; 27 – дополнительная; 28 – сошник; 29, 30 – ушные; I-V – жаберные дуги
Рис. 13. Череп и зубы судака
Рис. 14. Череп и зубы акулы
На рис. 15 представлены кости жаберной крышки.
Рис. 15. Кости жаберной крышки: 1 – предкрышка; 2 – межкрышка; 3 – крышка; 4 – подкрышка Кости жаберной крышки прикрывают пять жаберных дуг. На внутренней стороне четырёх дуг расположены жаберные тычинки, а на внешней - жаберные лепестки (органы дыхания). На пятой жаберной дуге лепестков нет. У некоторых рыб эта дуга превращается в нижнеглоточную кость, и на ней образуются глоточные зубы, которые достигают наибольшего развития у карповых. Они располагаются в 1…3 ряда. Форма и расположение глоточных зубов является систематическим признаком. При описании систематического положения рыбы определяют форму глоточных зубов. Для этого целиком извлекают нижнеглоточную кость и подсчитывают глоточные зубы на последней жаберной дуге. При записи число зубов с правой и левой стороны головы разделяют горизонтальной чертой. Если зубы расположены не в один ряд, то ближе к черте записывают количество зубов на нижнем (внешнем) крае кости, затем количество зубов в других рядах. Например, формула 5-6 означает, что зубы однорядные, на левой стороне их 6, а на правой – 5; 3.5-5.3 означает, что зубы двухрядные, на левой стороне в одном ряду 3, в другом 5 зубов, с правой стороны в одном ряду 5, в другом 3 зуба. У карповых имеется жерновок – роговое подушкообразное образование в верхней части глотки, служащее вместе с глоточными зубами для перетирания пищи. Шеи у рыб нет. Это обусловлено спецификой образа жизни и среды обитания – необходимостью разрезать головой воду, поэтому черепная коробка у взрослых рыб соединена с позвоночником неподвижно. У круглоротых отдельные хрящевые мозговые капсулы соединены с хордой, которая тянется от задней части черепа до хвоста в виде цельного несегментированного тяжа, состоящего из хрящевых и соединительных элементов (спинная струна), к которому сверху плотно прилегают хрящевые позвоночные дуги. Хорда осетровых также ещё не дифференцирована. У пластиножаберных (акуловых) рыб хорда покрыта хрящом и образует амфицельные (двояковогнутые) позвонки. Скелет грудного (плечевого) пояса состоит из трёх основных костей: ключицы, лопатки и коракоида. К нему прикрепляются грудные плавники. Он сочленён при помощи задневисочной кости с черепом. Скелет тазового поясов и плавников (парных и непарных). Тазовый пояс представлен у костистых рыб двумя срастающимися косточками, к которым прикрепляются лучи брюшных плавников. Он лежит в мышцах автономно, поэтому у некоторых видов может перемещаться далеко вперёд, даже на горло, а иногда и вовсе исчезать.
Скелет плавников являет собой опору, позволяющую рыбе пользоваться плавниками как рычагом или килем. У костистых рыб он представлен костными лучами, растягивающими кожистую плавательную перепонку. Различают лучи жёсткие и мягкие (нечленистые и членистые), которые, в свою очередь, разделяют на разветвлённые и неразветвлённые. Лучи соединяются с поясами конечностей или при помощи специальных косточек (грудной плавник), или непосредственно (брюшной плавник). Количество лучей в плавниках D и A подсчитывают при определении рыб, так как у костистых оно соответствует числу позвонков на определённом участке тела. Обычно определяют число лучей в спинном и анальном плавниках, а в случае описания нового вида – во всех плавниках. Жёсткие (неветвистые) обозначают римскими цифрами, мягкие (ветвистые) – арабскими. Например, D III 9 означает, что в спинном плавнике насчитывается 3 луча жёстких и 9 лучей мягких. В соответствии с формой и строением различают несколько типов хвостового плавника (рис. 16).
Рис. 16. Различные типы хвоста: а – симметричный; б – несимметричный (гетероцеркальный); в - ложносимметричный Все рыбы в эмбриональном и личиночном состоянии имеют прозрачную симметричную плавниковую кайму, огибающую конец позвоночника. Однако такая кайма у взрослых особей сохраняется только у круглоротых. У двоякодышащих позвоночник делит хвост на две равные части. Форма первично-симметричного хвоста (когда длинная ось тела делит его попалам) встречается редко. У древних групп рыб (акулы, осетровые) хвост несимметричный, неравнолопастный или гетероцеркальный: конец позвоночника резко загибается вверх, лопасти хвоста разные. У большинства костистых рыб, в том числе и у карпа, имеет место внутренняя асимметрия хвоста, образованная загнутым вверх уростилем, но при этом как вторичное явление развивается наружносимметричная его форма. Такая форма называется ложносимметричной. Функции осевого скелета и скелета поясов: опорная, кроме того, к ним прикрепляются двигательные мышцы. Пищеварительная система рыб Цель занятия: изучить пищеварительную систему рыб. Материалы: плакаты, таблицы. Пищеварительная система рыб подразделяется на: ротовую полость глотку пищевод желудок кишечник клоаку анус В пищеварительном тракте настоящих рыб различают ротовую полость, глотку, пищевод, желудок, кишечник (тонкая, толстая, прямая кишка заканчивающаяся анусом). У акул, скатов и некоторых других рыб перед анусом имеется клоака – расширение, куда изливается прямая кишка и протоки мочевой и половой систем. В строении различных отделов пищеварительного тракта имеется ряд особенностей табл. 15.
Таблица 15 Особенности строения отделов пищеварительного тракта рыб
Рис. 17. Пилорические придатки (указаны стрелкой) У рыб наблюдается зависимость длины пищеварительного тракта от рода пищи. Так, относительная длина кишечника (отношение длины кишечника к длине тела рыбы - L) составляет у растительноядных (пинагора и толстолобика) – 6…15, у всеядных (карася и карпа) – 2…3, у хищных (щуки, судака, окуня) – 0,6…1,2. Изучая пищеварительную систему рыб необходимо отдельно отметить две крупные железы: поджелудочную железу и печень. Обе железы носят общее название гепатопанцерес (hepatopancreas). Поджелудочная железа – сложная альвеолярная железа, производная кишечника, является компактным органом только у акул и немногих других рыб. У большинства рыб она визуально не обнаруживается, так как диффузно внедрена в ткань печени (большей частью), и поэтому её можно различить только на гистологических препаратах. Каждая долька связана с артерией, веной, нервным окончанием и протоком, выводящим секрет к желудочному пузырю. У карповых рыб (линь, серебряный карась, сазан) поджелудочная железа представлена скоплением специализированных групп клеток, которые локализуются в печени, брыжейке и жировой ткани кишечника, а также в селезёнке. В поджелудочной железе вырабатываются пищеварительные ферменты, действующие на белки, жиры и углеводы (трипсин, эрепсин, энтерококиназа, липаза, амилаза, мальтаза), которые выводятся в кишечник. У костистых рыб встречаются в паренхиме поджелудочной железы островки Лангерганса, в которых многочисленны клетки, синтезирующие инсулин, выделяемый прямо в кровь и регулирующий углеводный обмен. Таким образом, поджелудочная железа является железой внешней и внутренней секреции. Печень – крупная пищеварительная железа, по размерам уступающая у взрослых рыб только гонадам. Её масса составляет у акул 14…25 %, у костистых – 1…8 % массы тела. Это сложная трубчато-сетчатая железа, по происхождению связанная с кишечником. У зародышей является его слепым выростом. У большинства рыб, кроме некоторых лососевых, печень имеет многолопастную форму: в ней различают две, три, четыре, а у карповых даже семь лопастей. В печёночной паренхиме тянутся печёночные артерии, вены и желчные сосуды, собирающие желчь, вырабатываемую печёночными клетками. Желчные протоки проводят желчь в желчный пузырь (только у единичных видов его нет). Желчь благодаря щелочной реакции нейтрализует кислую реакцию желудочного сока. Она эмульгирует жиры, активирует липазу – фермент поджелудочной железы. Из пищеварительного тракта вся кровь медленно протекает через печень. В печёночных клетках кроме образования желчи происходит обезвреживание попавших с пищей чужеродных белков и ядов, откладывается гликоген, а у акул и тресковых (треска, налим и др.) - жир и витамины. Пройдя через печень, кровь по печёночной вене направляется к сердцу. Объём печёночных клеток меняется под влиянием интенсивности синтеза и расхода углеводов, которые обусловлены, в свою очередь, температурой окружающей среды, подвижностью, половой зрелостью рыб, интенсивностью питания и качеством пищи. Поэтому цвет и плотность ткани и общая масса печени сильно колеблются в зависимости от биологических особенностей рыбы и сезона года. При обильном полноценном питании печень приобретает красно-коричневый цвет с глянцевым отливом и некоторую упругость, масса её увеличивается; у голодающих рыб она становится дряблой, тусклой, мутной, желто-зелёной. Её объём и масса сильно уменьшаются. У прудовых карповых рыб к осени печень достигает максимальных размеров и массы и становится самой тяжёлой из всех органов в полости тела; к весне, после длительного зимнего голодания, масса её резко снижается. Сокращение объёма печёночных клеток после нереста установлено у радужной форели. Барьерная функция печени (очищение крови от вредных веществ) обусловливает её важнейшую роль не только в пищеварении, но и кровообращении.
Кровеносная система рыб Цель занятия: изучить кровеносную систему рыб. Материалы: плакаты, таблицы. Морфологическая и биохимическая характеристика крови различна у разных видов рыб в связи с систематическим положением, особенностями среды обитания и образа жизни. Внутри одного вида эти показатели колеблются в зависимости от сезона года, условий содержания, возраста, пола, состояния особей. Морфологическая и биохимическая характеристика крови разных видов рыб представлена в табл. 21. Таблица 21 Морфологическая и биохимическая характеристика крови разных видов рыб
Главным отличием кровеносной системы рыб от других позвоночных является наличие двухкамерного сердца (наполненного венозной кровью (за исключением двоякодышащих и кистепёрых)) и одного круга кровообращения. На рис. 23. представлена схема кровеносной системы костистой рыбы.
Рис. 23. Схема кровеносной системы костистой рыбы: 1 – венозная пазуха; 2 – предсердие; 3 – желудочек; 4 – луковица аорты; 5 – брюшная аорта; 6 - приносящие жаберные артерии; 7 – выносящие жаберные артерии; 8 – корни спинной аорты; 9 – передняя перемычка, соединяющая корни аорты; 10 – сонная артерия; 11 – спинная аорта; 12 – подключичная артерия; 13 - кишечная артерия; 14 – брыжеечная артерия; 15 – хвостовая артерия; 16 – хвостовая вена; 17 – воротные вены почек; 18 – задняя кардинальная вена; 19 – передняя кардинальная вена; 20 – подключичная вена; 21 – Кювьеров проток; 22 – воротная вена печени; 23 – печень; 24 – печёночная вена; чёрным показаны сосуды с венозной кровью, белым – с артериальной Сердце рыб состоит из одного желудочка и одного предсердия и помещается в околосердечной сумке, сразу за головой, позади последних жаберных дуг, то есть по сравнению с другими позвоночными сдвинуто вперёд. Перед предсердием имеется венозная пазуха, или венозный синус, со спадающими стенками; через эту пазуху кровь поступает в предсердие, а из него – в желудочек.
Расширенный начальный участок брюшной аорты у низших рыб (акулы, скаты, осетровые, двоякодышащие) образует сокращающийся артериальный конус. А у высших рыб – луковицу аорты, стенки которой сокращаться не могут. Обратному току крови препятствуют клапаны. Схема кровообращения в самом общем виде представлена следующим образом. Венозная кровь, заполняющая сердце, при сокращениях сильного мускульного желудочка через артериальную луковицу по брюшной аорте направляется вперёд и поднимается в жабры по приносящим жаберным артериям. У костистых рыб их четыре с каждой стороны головы – по числу жаберных дуг. В жаберных лепестках кровь проходит через капилляры и окисленная, обогащённая кислородом направляется по выносящим сосудам (их также четыре пары) в корни спинной аорты, которые затем сливаются в спинную аорту, идущую вдоль тела назад, под позвоночником. Соединение корней аорты спереди образует характерный для костистых рыб головной круг. Вперёд от корней аорты ответвляются сонные артерии. От спинной аорты идут артерии к внутренним органам и мускулатуре. В хвостовом отделе аорта переходит в хвостовую артерию. Во всех органах и тканях артерии распадаются на капилляры. Собирающие венозную кровь венозные капилляры впадают в вену, несущие кровь к сердцу. Хвостовая вена, начинающаяся в хвостовом отделе, войдя в полость тела, разделяется на воротные вены почек. В почках разветвления воротных вен образуют воротную систему, а выйдя из них, сливаются в парные задние кардинальные вены. В результате слияния вен задних кардинальных с передними кардинальными (ярёмными), собирающими кровь из головы, и подключичными, приносящими кровь из грудных плавников, образуются два Кювьерова протока, по которым кровь попадает в венозный синус. Кровь из пищеварительного тракта (желудка, кишечника) и селезёнки, идущая по нескольким венам, собирается в воротную вену печени, разветвления которой в печени образуют воротную систему. Собирающая кровь из печени печёночная вена впадает прямо в венозный синус. Кроветворение рыб по сравнению с высшими позвоночными отличается рядом особенностей табл. 22. Таблица 22 Особенности кроветворения рыб
Селезёнка рыб располагается в передней части полости т
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-12-30; просмотров: 532; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.147.27.152 (0.018 с.) |

 .
.


 Из мешкообразного выпячивания спинной части начала кишечника у рыб образуется плавательных пузырь – орган, свойственный только рыбам.
Из мешкообразного выпячивания спинной части начала кишечника у рыб образуется плавательных пузырь – орган, свойственный только рыбам.
 Рис. 15. Глоточные зубы сазана Рис. 16. Спиральный клапан (указан стрелкой)
Рис. 15. Глоточные зубы сазана Рис. 16. Спиральный клапан (указан стрелкой)


