Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Механизм действия гидрофильных гормонов
Гидрофильные гормоны построены из аминокислот, или являются производными аминокислот. Они депонируются в больших количествах в клетках желез внутренней секреции и поступают в кровь по мере необходимости. Большинство этих веществ переносятся в кровотоке без участия переносчиков. Гидрофильные гормоны не способны проходить через липофильную клеточную мембрану, поэтому действуют на клетки-мишени за счет связывания с рецептором на плазматической мембране.
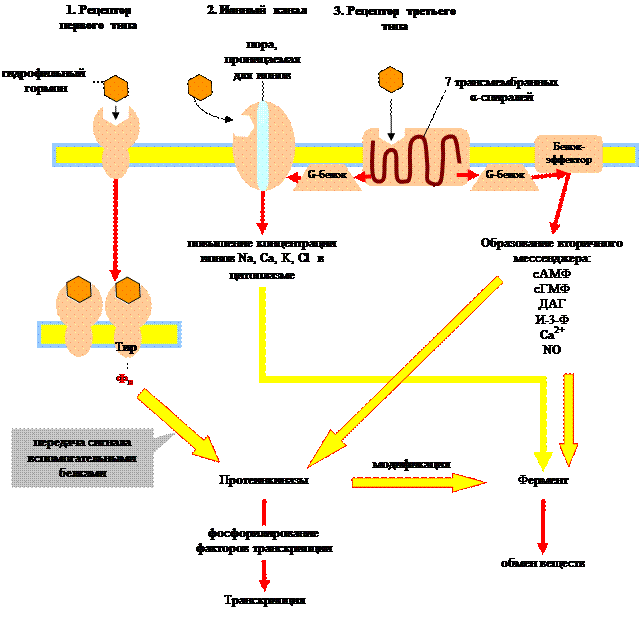
Рецепторы – это интегральные мембранные белки, которые связывают сигнальные вещества на внешней стороне мембраны и за счет изменения пространственной структуры генерируют новый сигнал на внутренней стороне мембраны.
Различают три типа рецепторов:
Связывание гидрофильного гормона с мембранным рецептором влечет за собой один из трех вариантов внутриклеточного ответа: 1) рецепторные тирозинкиназы активируют внутриклеточные протеинкиназы, 2) активация ионных каналов ведет к изменению концентрации ионов, 3) активация рецепторов, сопряженных с ГТФ-связывающими белками, запускает синтез веществ-посредников, вторичных мессенджеров. Все три системы передачи гормонального сигнала взаимосвязаны.
Рассмотрим преобразование сигнала G-белками, поскольку этот процесс играет ключевую роль в механизме действия целого ряда гормонов. G-белки переносят сигнал с рецептора третьего типа на белки-эффекторы. Они состоят из трех субъединиц: α, β и g. α-субъединица может связывать гуаниновые нуклеотиды (ГТФ, ГДФ). В неактивном состоянии G-белок связан с ГДФ. При связывании гормона с рецептором, последний меняет свою конформацию таким образом, что может связать G-белок. Соединение G-белка с рецептором приводит к обмену ГДФ на ГТФ. При этом происходит активация G-белка, он отделяется от рецептора и диссоциирует на α-субъединицу и β, g-комплекс. ГТФ-α-субъединица связывается с белками-эффекторами и изменяет их активность, в результате чего происходит синтез вторичных посредников (мессенджеров): цАМФ, цГМФ, диацилглицерин (ДАГ), инозит-1,4,5-трифосфат (И-3-Ф) и др. Медленный гидролиз связанного ГТФ до ГДФ переводит α-субъединицу в неактивное состояние и она вновь ассоциируется с β, g-комплексом, т.е. G-белок возвращается в исходное состояние.
Вторичные мессенджеры, или посредники, это внутриклеточные вещества, концентрация которых строго контролируется гормонами, нейромедиаторами и другими внеклеточными сигналами. Наиболее важными вторичными мессенджерами являются цАМФ, цГМФ, диацилглицерин (ДАГ), инозит-1,4,5-трифосфат (И-3-Ф), монооксид азота.
Механизм действия цАМФ. цАМФ является аллостерическим эффектором протеинкиназ А (ПК-А) и ионных каналов. В неактивном состоянии ПК-А является тетрамером, две каталитические субъединицы (К-субъединицы) которого ингибированы регуляторными субъединицами (R-субъединицы). При связывании цАМф R-субъединицы диссоциируют из комплекса и К-субъединицы активируются.
Активный фермент может фосфорилировать определенные остатки серина и треонина в более чем 100 различных белках и факторах транскрипции. В результате фосфорилирования изменяется функциональная активность этих белков.
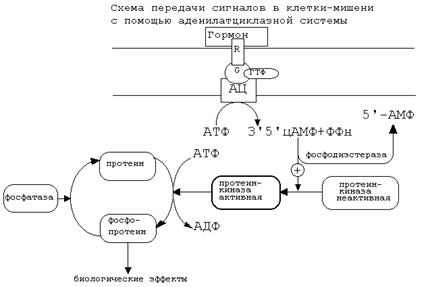
Если связать все воедино, то получается следующая схема аденилатциклазной системы:
Активация аденилатциклазной систтемы длится очень короткое время, потому что G-белок после связывания с аденилатциклазой начинает проявлять ГТФ-азную активность. После гидролиза ГТФ G-белок восстанавливает свою конформацию и перестает активировать аденилатциклазу. В результате прекращается реакция образования цАМФ.
Кроме участников аденилатциклазной системы в некоторых клетках-мишенях имеются белки-рецепторы, связанные с G-белками, которые приводят к торможению аденилатциклазы. При этом комплекс “GTP-G-белок” ингибирует аденилатциклазу.
Когда останавливается образование цАМФ, реакции фосфорилирования в клетке прекращаются не сразу: пока продолжают существовать молекулы цАМФ - будет продолжаться и процесс активации протеинкиназ. Для того, чтобы прекратить действие цАМФ, в клетках существует специальный фермент - фосфодиэстераза, который катализирует реакцию гидролиза 3',5'-цикло-АМФ до АМФ.
Некоторые вещества, обладающие ингибирующим действием на фосфодиэстеразу, (например, алкалоиды кофеин, теофиллин), способствуют сохранению и увеличению концентрации цикло-АМФ в клетке. Под действием этих веществ в организме продолжительность активации аденилатциклазной системы становится больше, то есть усиливается действие гормона.
Кроме аденилат-циклазной или гуанилатциклазной систем существует также механизм передачи информации внутри клетки-мишени с участием ионов кальция и инозитолтрифосфата. Инозитолтрифосфат - это вещество, которое является производным сложного липида - инозитфосфатида. Оно образуется в результате действия специального фермента - фосфолипазы “С”, который активируется в результате конформационных изменений внутриклеточного домена мембранного белка-рецептора.
Этот фермент гидролизует фосфоэфирную связь в молекуле фосфатидил-инозитол-4,5-бисфосфата и в результате образуются диацилглицерин и инозитолтрифосфат.
Известно, что образование диацилглицерина и инозитолтрифосфата приводит к увеличению концентрации ионизированного кальция внутри клетки. Это приводит к активации многих кальций-зависимых белков внутри клетки, в том числе активируются различные протеинкиназы. И здесь, как и при активации аденилатциклазной системы, одной из стадий передачи сигнала внутри клетки является фосфорилирование белков, которое в приводит к физиологическому ответу клетки на действие гормона.
В работе фосфоинозитидного механизма передачи сигналов в клетке-мишени принимает участие специальный кальций-связывающий белок - кальмодулин. Это низкомолекулярный белок (17 кДа), на 30% состоящий из отрицательно заряженных аминокислот (Глу, Асп) и поэтому способный активно связывать Са+2. Одна молекула кальмодулина имеет 4 кальций-связывающих участка. После взаимодействия с Са+2 происходят конформационные изменения молекулы кальмодулина и комплекс “Са+2-кальмодулин” становится способным регулировать активность (аллостерически угнетать или активировать) многие ферменты - аденилатциклазу, фосфодиэстеразу, Са+2,Мg+2-АТФазу и различные протеинкиназы.
В разных клетках при воздействии комплекса “Са+2-кальмодулин” на изоферменты одного и того же фермента (например, на аденилатциклазу разного типа) в одних случаях наблюдается активация, а в других - ингибирование реакции образования цАМФ. Такие различные эффекты происходят потому, что аллостерические центры изоферментов могут включать в себя различные радикалы аминокислот и их реакция на действие комплекса Са+2-кальмодулин будет отличаться.
Таким образом, в роли "вторых посредников" для передачи сигналов от гормонов в клетках-мишенях могут быть: - Циклические нуклеотиды (ц-АМФ и ц-ГМФ); - Ионы Са; - Комплекс “Са-кальмодулин”; - Диацилглицерин; - Инозитолтрифосфат
Механизмы передачи информации от гормонов внутри клеток-мишеней с помощью перечисленных посредников имеют общие черты: 1. одним из этапов передачи сигнала является фосфорилирование белков; 2. прекращение активации происходит в результате специальных механизмов, инициируемых самими участниками процессов, - существуют механизмы отрицательной обратной связи.
Гормоны являются основными гуморальными регуляторами физиологических функций организма, и в настоящее время хорошо известны их свойства, процессы биосинтеза и механизмы действия.
|
|||||||
|
|
Последнее изменение этой страницы: 2016-12-30; просмотров: 1285; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.137.178.133 (0.01 с.) |