Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Нейроанатомия функциональных состоянийСодержание книги
Поиск на нашем сайте
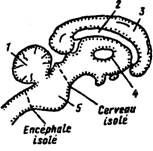
Открытие исследователем из Лос-Анжелеса Г. Мэгуном и итальянским ученым Дж. Моруцци в 1949 г. ретикулярной активирующей системы мозга имело решающее значение для понимания механизмов регуляции функциональных состояний. Они показали, что электрическая стимуляция срединной части ствола мозга (ретикулярной системы) во время сна животного пробуждает его и поддерживает бодрствование. По существу, изучение восходящей активирующей системы было начато еще раньше, уже классическими опытами Ф. Бремера с перерезками ствола мозга, результаты которых были опубликованы в 1935 г. Делая перерезки на уровне среднего мозга, он получил препарат, который назвал cerveau isolé (рис. 43). Его электрическая активность такая же, как у нормального спящего животного: в ЭЭГ постоянно возникают так называемые сонные веретена — регулярные высокоамплитудные колебания с частотой 8-12/с. При этом зрачок у животного сужен, что также типично для нормального сна. Другой препарат Ф. Бремер получил в результате рассечения ствола мозга на уровне его соединения со спинным (рис. 43). Препарат получил название encéphale isolé. Он имел ЭЭГ и реакции зрачка такие же, как и у нормального животного. Для него было характерно чередование «сна» и «бодрствования» (по показателям ЭЭГ и диаметра зрачка) (рис. 44). Оба препарата различались тем, что у cerveau isolé сенсорными раздражениями можно было вызвать лишь краткие периоды пробуждения, тогда как у encéphale isolé аналогичная стимуляция вызывала длительное пробуждение. Таким образом, у нормального животного и у encéphale isolé, по-видимому, существует центральный Рис. 43. Схема ствола мозга кошки, показывающая места перерезки при создании препаратов encéphale isolé и cerveau isolé Рис. 43. Схема ствола мозга кошки, показывающая места перерезки при создании препаратов encéphale isolé и cerveau isolé; 1 — мозжечок, 2 — мозолистое тело, 3 — кора, 4 — таламус, 5 — Варолиев мост
механизм поддержания бодрствования во время интервалов между раздражениями, но эта система отсутствует у препарата cerveau isolé. Именно этому центральному механизму в экспериментах Дж. Моруцци и Г. Мэгуна было найдено место. Их открытия хорошо объясняли эксперименты Бремера: источником ЭЭГ-активации являются не только сенсорные пути, но и ретикулярная формация среднего мозга. Животные с ретикулярными разрушениями становились сонными, обездвиженными и оставались такими по крайней мере в течение нескольких дней после Рис. 44. Сравнение ЭЭГ и величины зрачка у препаратов encéphale isolé и cerveau isolé Рис. 44. Сравнение ЭЭГ и величины зрачка у препаратов encéphale isolé и cerveau isolé. Быстрая и низкоамплитудная активность в ЭЭГ и расширенный зрачок, наблюдаемые у препаратов encéphale isolé, свойственны нормальному бодрствующему животному, а синхронизированные веретена в ЭЭГ и суженный зрачок типичны для cerveau isolé и нормального спящего животного (по Ф. Бремеру, 1937)
не активны, они редко разряжаются спайками. Но они первыми реагируют на наступление ПС. Их активность драматически возрастает и остается на этом уровне в течение всего периода ПС. Во время ПС кроме тонической активности у них можно видеть взрывы спайков, которые непосредственно предшествуют быстрым движениям глаз. По мнению Г. Шеперда [49], управление сном и бодрствованием осуществляется группой структур, которые образуют распределенную систему в мозге. Важнейшую роль в этой системе играют три стволовых центра, в каждом из которых действует особый медиатор. Состояние бодрствования связано с активностью норадренергических волокон (синее пятно), а глубокий медленный сон — с активностью серотонинергических волокон (дорзальные ядра шва). Интересную гипотезу, объясняющую взаимодействие этих центров в 1977 г. выдвинули исследователи из Гарвардского университета А. Хобсон и Р. Мак-Карли. Они считают, что чередование бодрствования и сна задается холинергическими гигантскими ретикулярными нейронами гигантоклеточного ядра моста, которые характеризуются самовозбуждающимися связями. Возбуждаясь, эти нейрону посылают импульсы к синему пятну и дорзальным ядрам шва. Во время бодрствования их активность подавлена за счет тормозных влияний из синего пятна. Под влиянием гигантоклеточного ядра моста находятся многие структуры мозга, которые и обусловливают различные тонические и фазические проявления ПС.
|
||||
|
|
Последнее изменение этой страницы: 2016-09-20; просмотров: 298; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.136 (0.034 с.) |