Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Сервоконтроль мышечных сокращенийСодержание книги
Поиск на нашем сайте
В связи с существованием непосредственного контроля со стороны центральной нервной системы над рецепторными процессами посредством гамма-системы афферентных волокон, нейронную организацию рефлекса нельзя больше рассматривать как дугу или простую последовательность «стимул — реакция». Существует множество данных, свидетельствующих о том, что и сами рецепторы контролируются центральной нервной системой — особенно те из них, которые воспринимают мышечные сокращения. Этот центральный контролирующий механизм действует как система обратных связей и процессов предвосхищения, что, по существу, и определяет корригирующие возможности сервомеханизма. Получено так много доказательств справедливости этого положения, что Р. Гранит, один из первых исследователей как моторных, так и сенсорных механизмов, вынужден был недавно заявить следующее: «... Что касается мотонейронов, то основные проблемы связаны не с рефлексами, хотя рефлексы всегда были и все еще остаются полезным средством анализа, необходимым в данной области знания. Основные проблемы касаются настройки или установки различных механизмов, посредством которых управляются мотонейроны — рефлекторно или как-либо иначе. Существует настройка мотонейронов с помощью нейромышечной интрафузальной системы гамма-волокон [находящейся в мышечных веретенах], а также посредством особых систем нейронных связей и нейрогормонов, действующих на альфа- и гамма-мотонейроны из высших уровней нервной системы» (Granit and Kellerth, 1967). Относительно сервоконтроля движений многое стало известно после эксперимента, проведенного в духе нейропсихологических исследований и давшего парадоксальные результаты. «Если мышца длительное время находится в состоянии сокращения под влиянием разрядов моторных импульсов, идущих от спинного мозга, удар тока, нанесенный на моторный нерв, приведет к возникновению судорожных всплесков на тензозаписи. Во время судорожного сокращения наблюдается прекращение разрядов моторных импульсов, идущих к мышце
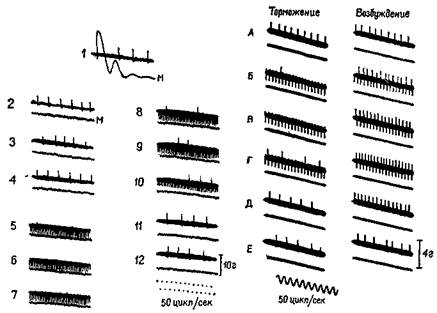
Рис. ХП-3. Периоды молчания в работе подошвенной мышцы человека, регистрация с помощью игольчатых электродов (нижняя запись). Под пяткой испытуемого закрепляется петля, и натяжение разгибателей щиколотки (верхняя запись) регистрируется при' давлении вниз на палец ноги. Время: 10 и 100 мсек; 5 суперпозиций в каждой записи; 1 — период молчания во время подергивания сухожилия, возникающего при ударе молоточком по ахиллесову сухожилию. При контакте молоточка с кожей возникает «всплеск» на кривой; 2 — запись во время рефлекса сокращения, вызванного электрическим раздражением афферентных волокон в подколенной ямке, 3 — запись во время судороги в латеральной части икроножной мышцы, вызванной стимуляцией выше этой мышцы. Обратите внимание, что подошвенная мышца сама по себе не возбуждается стимуляцией. Отметка времени (нижняя кривая на каждой записи) указывает 10 и 100 мсек (Granit, 1955). (рис. ХП-3). Это — период молчания. Это — рефлекторный акт (обсуждение этого вопроса см. Merton, 1951), который возникает, по-видимому, вследствие прекращения разрядов от мышечных веретен во время судороги. Именно эти разряды в обычных условиях вызывают рефлекс растяжения мышцы, и при их исчезновении мотонейроны замолкают. С точки зрения сервотеории, период молчания можно интерпретировать следующим образом: электрический ток размыкает замкнутую сервосистему и тем самым возбуждает поток добавочных моторных импульсов, который и вызывает сокращение мышцы; следовательно, механизм отрицательной обратной связи отключает спинальные моторные разряды до тех пор, пока мышца посредством расслабления не восстановит своей первоначальной длины. Надо отметить, что отрицательная обратная связь не только нейтрализует эффекты изменений нагрузки или интенсивности добавочного потока импульсов, но делает работу мышц независимой от утомления или изменения синаптической возбудимости, точно так же, как усилитель, сконструированный по принципу обратной связи, не чувствителен к изменениям интенсивности поступающего тока или к напряжению электронных ламп. Стабильность — вот единственное требование, предъявляемое к системе обратных связей: в усилителе это сеть обратных связей со стабильным сопротивлением, в мышцах же это неутомляемые сенсорные органы. Как известно из работ Мэтгьюза, мышечные веретена как раз и обладают этими свойствами... Возвращаясь к проприоцептивному механизму периода молчания, следует сказать, что мышечные, веретена прекращают свои разряды во время судороги потому, что они соединены параллельно с главными мышечными волокнами. При сокращении мышцы натяжение веретен уменьшается и они не посылают больше разрядов. Это «параллельное» соединение веретен и является ключом ко всей теории. Важная роль этого механизма состоит в том, что он дает возможность веретенам посылать сигнальную информацию о длине мышц, тогда как если бы веретена были соединены последовательно, они могли бы отвечать только на их напряжение. Таким механизмом, как серворефлекс растяжения, который столь очевидно обеспечивает поддержание определенной длины мышц, а не их напряжения, должны обязательно обладать рецепторы, сигнализирующие о длине мышц» (Merton, 1955, р. 248-259). Существуют, конечно, и другие рецепторно-эффекторные связи, контролирующие сокращения мышц. Есть рецепторы в сухожилиях и суставах — нельзя не учитывать также и растяжение кожи вокруг мышцы и суставов как источник информации для контроля над движениями. Хотя о, рамма-системе известно больше, чем о других механизмах контроля движений, иннервация, поступающая от мышечных веретен, является, по всей вероятности, основным механизмом, на котором надстраиваются другие контролирующие механизмы (рис. ХИ-4). Как было установлено, гамма-волокна бывают двух типов. Один тип реагирующих волокон обнаруживает постоянную частоту разрядов, если мышца сохраняет постоянную длину. Другой тип, напротив, обнаруживает уменьшение частоты разрядов во время завершения динамической фазы растяжения. Мэт-тьюз (1964) в обширном обзоре, посвященном этой теме, приходит к заключению, что такая двойная гамма-иннервация каждого мышечного веретена «обеспечивает относительно независимый контроль за общим режимом работы и отключением сервосистемы», поскольку каждый тип контроля раздельно управляется с помощью центральной стимуляции (рис. ХП-5). Этот анализ важен с двух точек зрения. Во-первых, он привлекает наше внимание к тому факту, что любое, даже самое простое мышечное сокращение должно управляться набором сигналов, которые адресуются либо только к мышечным рецепторам, либо идут одновременно с теми сигналами, которые поступают к сократительным элементам мышцы. Факты (см. следующие разделы) свидетельствуют о том, что некоторые области мозга, а именно базальные ганглии и передняя часть мозжечка, функционируют совместно, непосредственно управляя сократительной альфа-системой и опосредованно воздействуя на рецепторы, контролирующие гамма-разряды, благодаря чему и достигается подобная согласованность движений. Во-вторых, управление рефлекторным сервомеханизмом легко представить себе, поскольку этот процесс разыгрывается на
Время—>- Рис. XII-4. Примеры ответов сенсорных нервных волокон, идущих от сухожильного органа Гольджи (а) и рецептора мышечного веретена (б) и обеспечивающих растяжение и сокращение мышцы. Обратите внимание, что ответ рецептора мышечного веретена (который действует посредством у-системы) выражается как в форме растяжения, так и в форме удлинения мышцы. См. текст н р и с. ХП-5, где обсуждаются эти эффекты (Granit, 1955, см. Thompson, 1967). уровне рецепторов. Следовательно, нет необходимости в специальном устройстве типа клавиатуры фортепиано, с помощью которого импульсы, идущие от мозга, передавали бы мышцам сообщения; дело вовсе не в том, чтобы осуществить сначала одно, а потом другое сокращение, изменить длину того или иного из мышечных волокон. Существующее состояние сокращения мышечного волокна предопределяет вариативное состояние его рецепторов. Набор нервных импульсов взаимодействует с этим фоновым состоянием, изменяя режим работы мышечной сервосистемы и таким образом регулируя мышечное сокращение. Мозг все время должен посылать сигналы, организованные по пространственному и временному принципам, однако сообщения, передаваемые этими сигналами, не должны быть закодированы таким образом, чтобы прямо соответствовать сокращению или расслаблению мышцы, которое эти сообщения вызывают, поскольку эта информация уже содержится в вариативном состоянии рецептора. В следующей главе мы детально проанализируем эти сообщения, которые должны содержать подобный код. Подведем итоги. Нейрофизиологические данные свидетельствуют о существовании прямых анатомических афферентных путей, идущих от спинного мозга к мышечным рецепторам. Наиболее фундаментальным фактом является то, что основным
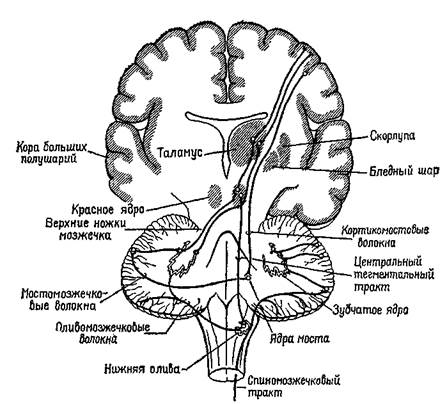
Рис. ХН-5. Двойная иннервация мышечного веретена с позиций концепции Т — О — Т — Е. принципом организации даже самого простого рефлекса является не дуга: стимул — центральная нервная система — реакция, а замкнутая сервосистема в виде последовательности: проба (test) — операция (operate)—проба (test)—результат (exit) (Т—О—Т—Е). Когда с помощью центральной нервной системы рефлексы объединяются в Сложные движения, эта интеграция не может быть эффективной при посылке серий сигналов непосредственно или исключительно сокращающимся мышцам, вызывая в них мелодии, как на клавиатуре фортепиано. Подобные сигналы только бы нарушали сервопроцессы. Чтобы предотвратить эти нарушения, наборы сигналов должны передаваться либо только мышечным рецепторам, либо одновременно с другими сигналами адресоваться непосредственно к мышечным волокнам. Таким образом, интегрированное движение в основном зависит от режима работы, настройки мышечных рецепторов. ПОДГОТОВКА К ОТВЕТУ Теория, излагаемая в этой книге, вполне согласуется с взглядами Ф. Меттлера (1967). Последний считает, во-первых, что «тонический фон движения служит своего рода канвой... на которой разыгрывается само движение». Этот фон находится под влиянием системы афферентных импульсов, которые отражают «потенциально доминирующие» мозговые процессы. Эта «канва», это фоновое состояние формируется благодаря работе ряда периферических сервомеханизмов, контролирующих мышечные сокращения. Далее, Меттлер признает существование «ассоциированных массивных движений (предмет данного раздела), на фоне которых вырисовываются отдельные детали...». Массивные процессы регуляции мышечного тонуса осуществляются базальными ганглиями, самыми передними структурами ствола мозга. Наконец, «на этом фоне статических и глобально действующих сил кора мозга ответственна за точность и интенсивность движений, з также за изменение скорости уже начавшегося движения и той скорости, с которой это движение усиливается и прекращается». Статическое состояние функций мышцы называется ее тонусом. Однако тонус — это не величина спастичности или вялости сократительной ткани. Это, скорее, состояние готовности всего нейромышечного аппарата к действию, предварительное условие всякого действия. Центральными структурами с замкнутыми системами связей, участвующими в регуляции тонической мышечной активности, являются базальные ганглии (рис. XI1-6) и передняя часть мозжечка (рис. ХП-10). Поражение этих структур заметно изменяет состояние готовности мышечного аппарата: так, удаление переднего отдела мозжечка ведет к вялости, потере мышечного тонуса, а удаление базальных ганглиев — к полной обездвиженности, невыразительности мимики, маске Паркинсона и тремору конечностей, когда последние находятся в состоянии покоя. Филогенетически эти структуры мозга развивались, когда позвоночные впервые приспособились к передвижению по земле. Таким образом, эти структуры интимно участвуют в механизмах, регуляции позы, создающих то состояние готовности к ответу, тот фон, на котором может начаться дискретное действие. Регуляция мышечной активности с помощью замкнутых кольцевых связей базальных ганглиев и передних отделов мозжечка была предметом большого числа исследований, однако в этом вопросе до сих пор нет полной ясности. По-видимому, в этих процессах регуляции участвует прежде всего гамма-система, хотя, как уже говорилось выше, альфа- и гамма-влияния участвуют также в интеграции движений. Наиболее существен-
Рис. ХП-6. Расположение базальных ганглиев внутри больших полушарий головного мозга (Krieg, 1966). ные характеристики системы в значительной мере зависят от количественного и временного соотношения тех реципрокных взаимосвязей, которые установились между альфа- и гамма-активностью, а также от пространственной координации повторяющихся наборов сигналов в данной системе (рис. XI1-7 и ХН-8). Эти характеристики трудно определить в экспериментальных ситуациях, в которых обычно применяются анестезирующие средства, что делает бесполезными попытки воспроизвести естественные механизмы регуляции позы. Тем не менее современные знания убеждают в том, что модуляция состояний готовности в мышечном аппарате осуществляется сервосистемой «базальные ганглии — передние отделы мозжечка» прежде всего посредством угашения непрерывных колебаний, присущих периферическому сервомеханизму с отрицательными обратными связями — механизму, определяющему рефлекторный акт. Это угашс-ние колебаний, вероятно, возникает благодаря пространственной координации, торможению и совместному действию реципрокных аппаратов различных сервомеханизмов. С помощью этих механизмов тоническое фоновое состояние моторной системы приобретает изменчивость и гибкость. ФАКТ ПРОИЗВОЛЬНОГО ДВИЖЕНИЯ Сервоконтроль имеет и другой аспект. Иногда сервомеханизм не только регулирует движение «на выходе», но и создает предварительную установку к следующему этапу, то есть
Рис. XI1-7. Торможение разряда мышечного веретена при раздражении передней доли мозжечка. Верхние кривые на каждой записи показывают разряд веретена, распространяющийся вверх от черты, и артефакты от применения стимула, которые направлены вниз. Нижние кривые в каждой записи — миограммы. Децеребрированное животное. Икроножная мышца. Начальное давление — 66 гр. Миограмма записана при максимальном усилении (см. запись 12), за исключением 1-й записи, в которой сокращение (клони-ческое) в ответ на единичное раздражение электрическим током икроножного нерва выявляет молчащий период; 2—4 — контрольные записи перед стимуляцией; 5—10 — во время стимуляции мозжечка током 140 имп/сек при длительности импульса — 1 мсек, в течение 26 мсек; 5—7 — через 18—20 сек; 8—10 — непосредственно после прекращения стимуляции. Обратите внимание на падение частоты разрядов веретена от 20 имп/сек до нерегулярных разрядов порядка 5 имп/сек (Granit, Kaada, 1952; Granit, 1955), Рис. ХИ-8. Записи активности веретена подошвенной мышцы. Влияние стимуляции тормозных и активирующих участков ба-зальных ганглиев и других структур ствола мозга. Как и на рис. ХП-7, верхняя линия иа каждой записи показывает разряд веретена (подъем линий вверх) и артефакт от стимуляции (опускание линий вниз). А — средняя линия; В — первая запись во время стимуляции (обратите внимание на артефакты при электрическом раздражении); В — во время стимуляции; Г — последняя запись перед прекращением стимуляции; Д, Е — сразу же после прекращения стимуляции. На нижней кривой — миограмма. Начальное давление — 55 гр. Кошка под дозированным хлоролозовым наркозом (Eldred, Granit, Merton, 1953; Granit, 1955), обеспечивает то предшествующее двигательной активности состояние, которое нуждается в стабилизирующих влияниях сервомеханизма. Эти процессы предвосхищения (рис. ХП-5) широко распространены в моторной системе и обозначаются как произвольные, волевые или преднамеренные движения и действия. Современный уровень разработки проблем, связанных с произвольной активностью, еще не дает возможности понять механизмы нервного контроля поведения; хорошим началом в этом отношении могут служить исследования гамма-системы. Приведем еще одно-высказывание Мертона. «Что же происходит во время произвольного сокращения? Легко осуществить произвольное сокращение мышц при устойчивом уровне двигательной активности; эти движения совершаются даже быстрее, чем самые быстрые судорожные сокращения. Скорость возникновения небольшого судорожного сокращения мышц лишь в малой степени соответствует скорости произвольного усиления. Здесь возникает очень интересная проблема: если во время такого усиленного сокращения мышц сервомеханизм будет продолжать действовать так же, как прежде, то он явно будет способствовать противоположному эффекту. Сразу же после начала произвольного сокращения мышечное веретено будет укорачиваться и возникающее в результате этого прекращение возбуждающих афферентных импульсов будет интенсивно подавлять мотонейроны... как если бы сокращение мышцы вызывалось небольшой стимуляцией двигательного нерва. Это значит, что дополнительные моторные разряды, возникающие при возбуждении мотонейронов спинного мозга, должны вызывать гот же самый эффект в замкнутой сервосистеме, что и импульсы, возникающие вокруг этой системы в их аксонах под влиянием электрического тока. В любом случае сервомеханизм должен энергично действовать, чтобы прекратить эти механические эффекты. Если эти выводы из экспериментов верны, то это означает, что во время произвольного сокращения следует преодолеть или устранить сопротивление, оказываемое сервосистемой... необходимость возбуждения основных мотонейронов, связанных с интрафузальными мышечными веретенами посредством нисходчщих импульсов, идущих из спинного мозга, исчезает. Возбуждение целиком поступает к небольшим мотонейронам (рецепторам мышечных веретен), н сервомеханизм запускает затем в действие основные мышцы. Таким образом, в произвольном сокращении мышцы участвует тот самый механизм, который обеспечивает постоянное фоновое сокращение, а именно: сервомеханизм, поддерживающий в мышце такой же ритм, как и в ее веретенах; следовательно, мышца находится под точно таким же стабилизирующим влиянием обратных связей, как и мышечные веретена. Предположение, что возбуждение поступает по основным мотонейропам, явно ошибочно, поскольку именно этот тип вмешательства, замкнутая система обратных связей и должна нейтрализовать. Замкнутая система наилучшим образом активируется в результате изменения режима работы ее «гасящих» детекторов (null-detektor). Эта гипотеза в известной мере подтверждается экспериментами, недавно проведенными на нервно-мышечных приборах, которыми снабжена грудпая мышца. Хант (1951), в частности, наблюдал во время рефлекторного сокращения этой мышцы разряды моторных импульсов, посылаемых к интрафузальным мышцам (рецепторам веретен), предвосхищающие появление активности в основных мотонейронах» (Merton, 1953, р. 251—253). Таким образом, произвольные движения, как правило, начинаются с активации интрафу зальных мышечных веретен. Исключением из этого правила является тот случай, когда движения возникают неожиданно. Вследствие сравнительно малого диаметра гамма-волокон и большой длины всего пути скорость проведения слишком мала, чтобы объяснить внезапные мышечные сокращения. Однако подобные движения относительно просты и гамма-система быстро включается в контроль над этими движениями. Следовательно, нельзя полностью игнорировать прямой путь к основным мотонейронам. Ограниченный объем предварительной информации, которая программирует процесс «сокращения — расслабления», может быть передан по этим путям, если интервал между началом и окончанием процесса будет достаточен для того, чтобы могла включиться в действие гамма-сервосистема. Работы Дж. Бермана и его сотрудников (Taub, Bacon and Berman, 1965) установили как верхнюю, так и нижнюю границы такого рода предпрограммирования движений. Берман использовал в эксперименте обезьяну, у которой спинной мозг ла всем его протяжении был полностью деаффектирован посредством удаления дорсальных корешков. Эта операция вызывает нарушение замкнутого сервомеханизма, обслуживающего движения. Однако после деафферентации у этих обезьян все же сохранялись условные реакции (сгибания конечностей). У человека же подобная деафферентация, широко практикуемая в настоящее время, чтобы снять боли и другие сенсорные нарушения, приводит к серьезным расстройствам автоматизированных движений, выполняемых соответствующей конечностью. Итак, управление рефлекторными и интегрированными произвольными движениями независимо от того, вызываются ли они внешними или внутренними факторами, осуществляется в основном посредством настройки рецепторов мышечных веретен, включенных в гамма-сервосистему. Центральный контроль над движениями сводится, следовательно, к центральному контролю за рецепторными процессами. Эту проблему мы уже обсуждали во второй части, более подробно рассмотрим ее в четвертой части книги. Теперь мы сформулируем парадокс, который не укладывается в рамки узкого бихевиоризма: даже относительно простые, доступные для наблюдения двигательные механизмы связаны с рецепторными процессами, регулируемыми центральной нервной системой. БЫСТРОДЕЙСТВУЮЩИЕ МЕХАНИЗМЫ ПРЕДВОСХИЩЕНИЯ Признав факт существования произвольных движений, мы должны ответить на вопрос: как же они осуществляются? Ответ на этот вопрос можно дать на основе анализа процессов предвосхищения, с одной стороны, и раскрытия сущности предпро- 25S Рис. XI1-9. Срединное сеченне через полушария и ствол мозга, на котором видно располошение и главные связи мозжечка Модифицированная схема Крига (1966). граммирующих процессов, подготавливающих результаты ряда движений прежде, чем они будут реализованы, — с другой. Вспомним, что в гл. V указывалось, что для процессов предвосхищения необходим только приблизительный план, черновой набросок тех операций, которые должны быть выполнены. Как только будет выработан такой приблизительный план движений, дальнейшую их отшлифовку может взять на себя обычная сервосистема обратных связей. На основании нейро-анатомических и нейрофизиологических данных можно допустить, что некоторые аппараты нервной системы идеально приспособлены для обеспечения этой общей планирующей функции; и, как показывают результаты поведенческих экспериментов, эти функции действительно осуществляются такими аппаратами. Центральным звеном этой управляющей системы являются филогенетически новейшая часть мозжечка, эта удивительная
структура, панорама которой открывается взору лишь при определенном сечении (рис. XII-9, ХН-10). Как уже отмечалось выше, передняя часть мозжечка развилась при переходе морских позвоночных к наземному образу жизни. До этого момента мозжечок функционировал главным образом как дополнительный вестибулярный контролирующий механизм. Рыбы поддерживают ориентацию в соответствии Рис. ХН-10. Схематическое изображение борозд и долек мозжечка (Lar-sell, 1951; Jansen, Brodal, 1958; Angevine et al., 1961). Части мозжечка, расположенные сзади по отношению к постлатеральной борозде, составляют клочок (archicerebellum), а части мозжечка, расположенные спереди от первичной борозды, составляют переднюю дольку (paleocerebellum). Neoce-rebellum расположен между первичной и постлатеральной бороздами. Римские цифры обозначают только разделы червя мозжечка (Truex, Carpenter, 1969). с осью своего тела; их движения начинаются от этой основной линии. Амфибия и другие земноводные имеют дополнительную основную линию, это земля под ними, «низ», определяемый посредством гравитационных сил. Поэтому для организма становятся полезными новые вертикальные ориентиры позы и вследствие этого развивается передняя часть мозжечка. Наконец, приматы в известной мере освобождаются от гравитационной основной линии благодаря развитию двигательных навыков и приспособляются к древесному образу жизни и прямой позе. Теперь полушария мозжечка становятся доминирующими. У человека повреждение этих отделов мозга ведет к нарушению координации движений, к грубым промахам при слежении за целью и к атаксии при выполнении произвольных движений.
Рис. XII-11. Схематическое изображение коры мозжечка в сагиттальной и поперечной плоскостях, показывающее расположение клеток и волокон. •Ср. с рис. XII-12, где дано изображение тех же клеток и волокон в поперечном разрезе (Truex, Carpenter, 1969). К счастью для нейрофизиологии, полушария мозжечка имеют удивительно гомогенное строение (рис. XII-11) и поэтому относительно просты для анализа. Их основными связями являются: 1) связи через спинной мозг со структурами, обеспечивающими афферентацию движений; 2) связи с корой больших полушарий (рис. ХП-9). Благодаря этим связям полушария мозжечка могут ■функционировать, как компаратор, сопоставляющий сигналы, поступающие от коры больших полушарий, с сигналами, идущими ■от периферии. Сущность этих процессов сравнения становится особенно интересной в свете последних детальных анатомических и физиологических данных (рис. XII-12). Полушария мозжечка.имеют кору, которая представляет собой как бы «прямоугольную решетку», напоминающую по своей конфигурации вертикальные ■колонки, обнаруженные в коре больших полушарий. Правда, в отличие от коры все эти мозжечковые решетчатые структуры
«фактически идентичны. Решетка построена таким образом, что, минуя один или два синапса, все поступающие в мозжечок импульсы преобразуются в тормозные сигналы. Этот невероятно развитый тормозной механизм должен служить следующим целям: «Это исключительное превращение всех поступающих сигналов в тормозные посредством самое большее двух синаптических реле придает мозжечку с точки зрения воздействия на входной сигнал характер аппарата, заглушающего колебания в системе. Вследствие этого становится невозможным непрерывное хранение информации посредством импульсов, циркулирующих по сложным нейрональным путям, как ото имеет место в коре больших полушарий или на путях между корой и базальными ганглиями. Через 30 мсек после поступления сигнала в кору мозжечка вызванные разряды уже не регистрируются. ... остальные отделы центральной нервной системы требуют от мозжечка, по-видимому, не осуществления сложных реверберационных циклов в пределах самого мозжечка, а быстрой и четкой реакции на поступление какого-либо частного вида информации» (Eccles, Ito, Szentagothai, 1967, p. 311). Быстрое стирание содержания записи предполагает, что последовательность операций сличения выполняется тоже довольно быстро. Эта особенность системы выходит за рамки обычного сервомеханизма обратных связей, в котором для длительного поддержания устойчивого состояния требуется постоянная информация о результатах движения. Т. Рач в своей работе, опубликованной в «Экспериментальной психологии» (1951), правильно сформулировал эту проблему, причем значительно раньше, чем были собраны соответствующие факты. «Процесс циркуляции возбуждения по замкнутому кругу от коры больших полушарий к [нео]-мозжечковым структурам выполняет функцию не столько устройства, корригирующего ошибки, сколько составляет часть Рис. ХП-12. Схема, показывающая распространение тормозных полей (затемненные области) в случае возбуждения узкого пучка параллельных волокон. Верхняя часть рисунка — поперечный разрез листа червя мозжечка; нижняя часть — вид на поверхность листа червя сверху. Предполагается, что существует «минимально эффективный пучок» одновременно возбужденных параллельных волокон, который, по-видимому, способен стимулировать организованные в ряды клетки Пуркинье, звездчатые и корзинчатые клетки. Клетки Гольджи, имеющие самую развитую сеть дендри-тов, вероятно, наиболее приспособлены к возбуждению большими группами одновременно возбужденных параллельных волокон. Следовательно, эффективная стимуляция клеток Гольджи — как отрицательная обратная связь — может перекрыть всю «входную информацию», поступающую по мшистым волокнам. Таким образом, систему клеток Гольджи можно рассматривать как «фокусирующее» устройство, которое ограничивает — или усиливает — активность гранулярных нейронов (через систему параллельных волокон) в относительно узких группах (Eccles, Ito and Szentagothai, 1967). механизма, который может мгновенно осуществлять предвосхищение влияния. Подобный замкнутый контур, хотя и не информированный о конечных результатах движения, мог бы, как говорится, «вчерне» программировать движение и таким образом сократить досадные издержки, связанные с коррекцией движения посредством аппарата обратных связей, который использует поступающую извне информацию» (Ruch, 1951, р. 205). Основная функция мозжечка, связанная с объемом регистрации, заключается в том, чтобы составить схему движения без детальной его разработки. В действительности такое кратковременное состояние, которое аналогично тому, что создается порой больших полушарий, формируется на фоне спонтанной активности. «Таким образом, следует предположить, что даже в условиях минимального сенсорного «входа» существует состояние динамического равновесия в уровне активности различных типов нейронов мозжечка.. Отрицательный образ обобщенного результата работы коры мозжечка формируется в ходе процесса, аналогичного созданию скульптуры из камня. Форма, обладающая пространственно-временными признаками, достигается постепенно посредством набора тормозных импульсов, подавляющих «бесформенные» фоновые разряды нейронов, расположенных на более низких уровнях, подобно тому как вечные формы в скульптуре создаются путем очень избирательного удаления резцом всего лишнего из первоначально бесформенного камня» (Eccles, Ito, Szentogothai, 1967). Подробнее я остановлюсь на этом приборе и на его аналоге-в коре больших полушарий в гл. XIII. Здесь же мы должны только констатировать, что механизмы предвосхищающего' схематичного планирования движения можно рассматривать в качестве эквивалента быстродействующего компьютера, оценивающего конечный результат последовательной серии движений. Сходство между голограммой и процессом формирования образа заставляет предположить, что здесь имеет место процесс типа преобразования Фурье, однако и об этом речь будет идти в следующей главе. Во всяком случае, результаты предварительного расчета движений всегда немедленно поступают как к периферической мускулатуре, так и в моторную область коры больших полушарий через различные мозжечковые пути. С инженерной точки зрения подобный быстрый расчет ожидаемого «выхода» в настоящее время считается исключительно полезным приспособлением (Kelley, 1968). По-видимому, система «мозг—поведение» уже давно овладела им. РЕЗЮМЕ Проблема мозговой организации поведения во многих отношениях является обратной стороной формирования Образа (восприятий и чувств). Вследствие повсеместного существования в нервной системе механизма обратной связи (и механизма предвосхищения) контроль над движениями достигается не не- го'* посредственно с помощью сигналов, адресующихся к сокращающимся мышечным волокнам, а посредством сигналов, адресующихся к рецепторам, которые регулируют мышечное сокращение. Таким образом, мышечные рецепторы становятся частью гармоничной системы (сервомеханизма Т—О—Т—Е), назначением которой является регуляция мышечных реакций в ответ яа изменения внешней среды. Короче говоря, нервная регуляция поведения осуществляется посредством воздействия на ре-щепторные процессы, а не через непосредственный контроль мышечных сокращений. Глава XIII ДЕЙСТВИЯ МОТОРНАЯ КОРА Четкие, избирательные движения, лежащие в основе поведения, осуществляются моторной корой. Эта кора, расположенная в прецентральной извилине полушарий (рис. XIII-1), связана с базальными ганглиями и мозжечком. Каковы же функции этой моторной коры? Уже на протяжении целого столетия идет горячий спор об особенностях организации центральной моторной системы. Некоторые авторы (Woolsey, Chang, 1948) придерживаются точки зрения, согласно которой в моторной зоне существует анатомическое «точка в точку» представительство мышц и даже волокон и моторная кора играет роль клавиатуры для всякой другой церебральной активности и прежде всего для всех произвольных действий. Напротив, другие авторы (например, Phillips, 1965) указывают, что рецептивные поля соседних корковых нейронов охватывают широкий набор мышц, хотя большинство активных мышц, зарегистрированных при раздражении какого-либо одного участка коры, связано лишь с определенным суставом. С таким представлением согласуется тот факт, что электрическое раздражение моторной коры вызывает движения, интегрированные в определенную последовательность мышечных сокращений, и что движения, вызываемые отдельным возбуждением, частично зависят от состояния мозга и положения конечностей, что и отражается на их реакциях при стимуляции. Эти нейрофизиологические данные всегда интерпретировались таким образом, что в моторной зоне коры представлены движения, а не мышцы. Несколько лет назад я еще раз вернулся к этому спору, повторил многие критические эксперименты и накопил новые наблюдения с помощью некоторых дополнительных технических устройств (Pribram, et al., 1955—1956). Результаты этих экспериментов и наблюдений показывают, что основная роль моторных зон коры заключается не в контроле за индивидуальными мышцами или специфическими движениями. Моторная кора, скорее, играет более важную роль в организации.
Рис. XIII-1. Представительство моторики в прецентральной коре человека, в — схема поверхности левого полушария мозга, показывающая расположение прецентральной извилины; о — увеличенная прецентральная извилина, видимая в том же ракурсе, что и на рис. в; б — медиальная поверхность прецентральной извилины. В действительности у различных людей наблюдается значительное перекрытие зон и их вариативность, однако в целом эта схема сохраняется. Схема составлена на основе данных Форстера, Пенфилда, Болдри и др. по электрической стимуляции лрецентральной коры и наблюдений за возникающими при этом движениями (Krieg, 1966). Рис. XIII-2. А — фотография поперечного сечения мозга через; таламус, гитлаламус и базальные ганглии на уровне зрительной хиазмы. Гб означает гипоталамическую борозду в стенке третьего-желудочка. ВПМ и ВПЛ означают вентральное постмедиальное и вентральное постлатеральное ядра таламуса. Методика окрашивания миелина по Вейгерту; Б — фотография поперечного сечения самых верхних отделов спинного мозга месячного ребенка. Методика окрашивания миелина по Вейгерту. Отметьте сходство в расположении и конфигурации задних рогов спинного мозга и задних отделов таламуса, а также передних рогов спинного мозга и базальных ганглиев (и субталамуса) (Truex, Carpenter, 19(39)..
действия — действия, определяемого не с точки зрения участвующих в нем мышц, а с точки зрения обеспечения внешнего выражения психологической установки или плана. Это навело меня на мысль, что центральный моторный механизм является в некотором смысле подобием сенсорной системы и что поражение моторной коры вызывает «скотомы в действиях» по аналогии со скотомами в зрительном поле, возникающими при поражении зрительной коры. Появляется все больше данных в пользу этой точки зрения. Вначале я столкнулся со следующим анатомическим парадоксом. Клетка моторной коры получает импульсы через д
|
||||
|
|
Последнее изменение этой страницы: 2016-09-20; просмотров: 328; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.20 (0.022 с.) |