Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Эфферентный контроль входа — петля обратной связиСодержание книги
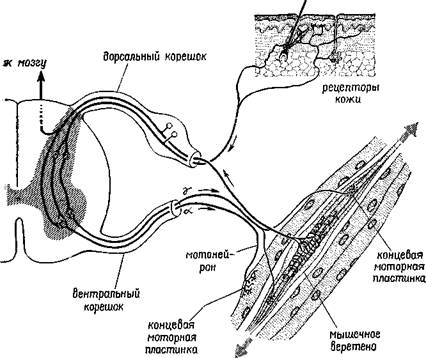
Поиск на нашем сайте Пересмотр концепции рефлекторной дуги стал необходим из-за новых экспериментальных данных, которыми не располагал Шер-рингтон. Эти данные показывают, что все входные системы организма непосредственно контролируются центральной нервной системой. Таким образом, выходные волокна — эфференты — регулируют не только движения организма, но и его рецепторы» а следовательно, и его сенсорные функции. Такой пересмотр классического понятия рефлекторной дуги начался с экспериментов, которые продолжали анализ функций! нервных волокон, образующих вентральный корешок (см. рис. V-2). Анатомический анализ показал, что одна треть этих волокон — группа волокон с маленьким диаметром, названная ■у-волокнами (в отличие от а- и р-волокон — большого диаметра)», оканчивается не в самой ткани сокращающихся мышц, а в особых рецепторах, названных мышечными веретенами. Эти веретена получают информацию о величине напряжения мышечных волокон, внутри которых они находятся. То, что эти веретена» иннервирует такое большое число волокон центрального корешка, весьма озадачило физиологов, разделявших концепцию рефлекторной дуги. Были проведены новые эксперименты, чтобы найти решение этой проблемы. Наиболее поразительные результаты, были получены при электрическом раздражении дистального (обращенного к мышце) конца перерезанного вентрального корешка*
Рис. V-2. Общая схема системы гамыа-мотоиейроиа (см Thompson, 1967). у которого были удалены путем рассечения волокна большого диаметра. Оказалось, что такая стимуляция не вызывает непосредственно сокращения мышц и, следовательно, у-волокна не являются зфферентами, а их функция — моторной. Еще более интересные результаты были получены во время стимуляции Y-эфферентных волокон и записи электрической активности дорсального корешка. В результате стимуляции вентрального корешка оказалась заметно сниженной спонтанная активность в афферентах дорсального корешка (Granit, 1944). Таким образом, Y-система образует отрицательную обратную связь, регулирующую вход, она берет свое начало от мышечных веретен, действуя в значительной степени так же, как механизмы адаптации и привыкания, описанные в гл. III. Далее было установлено, что активность афферентов мышечных веретен испытывает влияние через у-эфферентные связи со стороны спинного мозга (клеток малого вентрального рога), ствола мозга, мозжечка и даже коры (Kuffler .and Hunt, 1952; Leskell, 1945). Как будет подробно рассмотрено в гл. XII и XIII, эти экспериментальные результаты заставляют предположить, что регуляция моторных функций организма, его-поведения осуществляется скорее посредством особых рецепторов, которые тесно связаны с движением, а не в результате прямого управления мышечными сокращениями. Поэтому мы отложим дальнейшее изложение наших соображений по поводу нейронного контроля поведения до тех пор, пока не проанализируем в деталях тот механизм, с помощью которого осуществляется организация происходящих в рецепторах процессов. Вначале думали, что моторная система уникальна и центральный контроль над рецепторами существует только в двигательной
[20мк& 20мсек Рис. V-3. Запись электрических ответов на звуковой щелчок с помощью биполярных электродов, вживленных в зрительный нерв. Запись осуществлена непосредственно на фотопленку с осциллоскопа методом суперпозиции. Левая запись была сделана, когда животное обнаруживало реакцию внимания, а правая — во время его отвлечения (Spinelli, Pribram and Weingarten, 1965). системе. Однако очень скоро стали накапливаться данные о том, что кожные рецепторы (Hagbarth and Kerr, 1954), слуховой афферентный механизм (Galambos, 1956; Rasmussen, 1946; Desmedtr 1960; Dewson, 1968) и обонятельная чувствительность (Kerr and Hagbarth, 1955) также являются объектом прямой регуляции со< стороны центральной нервной системы. Не сразу удалось доказать существование эфферентного контроля зрительного входа; первые исследования (Granit, 1955; Hernandez-Peon and Schererr 1955) были подвергнуты критике потому, что еще не найдены морфологически эфферентные волокна в сетчатке. Поэтому в моей лаборатории были проведены исследования в надежде, что эфферентный контроль над зрительным входом станет вполне реальным фактом. В этих исследованиях были использованы только что появившиеся компьютеры и микроэлектродная техника. Результаты опытов показали, что незрительные стимулы (звуковые щелчки и удары по лапе) вызывают ответы в зрительном нерве у кошек (мышцы которых, в том числе и мышцьь зрачка, были обездвижены). Кроме того, электрическая активность -сетчатки (измеряемая с помощью электроретинограммы) и аффе-рентов, берущих начало в сетчатке, также меняется в результате такой незрительной стимуляции (Spinelli, Pribram and Weingar-ten, рис. V-3). Наконец, как и в случае других сенсорных" модальностей, стимуляция соответствующей части коры головного мозга вызывала изменение в рецептивных полях ганглиозных клеток сетчатки, например их величины (Spinelli and Pribram, 1967). Результаты этих экспериментов убеждают в том, что организация зрительного механизма сходна с организацией других сенсорных -систем, в которых существует центральный контроль над сенсор-лым выходом.
|
||
|
|
Последнее изменение этой страницы: 2016-09-20; просмотров: 311; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.102 (0.008 с.) |