
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
II. Условия выполнения законов МенделяСодержание книги
Поиск на нашем сайте
Приложение к генетике I. Механизмы, лежащие в основе законов Г. Менделя: 1. Мейоз 2. Оплодотворение II. Условия выполнения законов Менделя Законы И. Менделя являются фундаментальными законами генетики (подобно законам Ньютона в физике). Однако они (как и любые законы природы) выполняются только при наличии определенных условий:
При Несоблюдении перечисленных условий наследование признаков приобретает более сложный характер. III. Условия выполнения закона расщепления при моногибридном скрещивании Расщепление 3: 1 по фенотипу и 1: 2: 1 по генотипу выполняется приближенно и лишь при следующих условиях: 1. Изучается большое число скрещиваний (большое число потомков). 2. Гаметы, содержащие аллели А и а, образуются в равном числе (обладают равной жизнеспособностью). 3. Нет избирательного оплодотворения: гаметы, содержащие любой аллель, сливаются друг с другом с равной вероятностью. 4. Зиготы (зародыши) с разными генотипами одинаково жизнеспособны. IY. Условия выполнения закона независимого наследования 1. Все условия, необходимые для выполнения закона расщепления. 2. Расположение генов, отвечающих за изучаемые признаки, в разных парах хромосом (несцепленность). 4. Условия выполнения закона чистоты гамет: 1. Нормальный ход мейоза. 2. В результате нерасхождения хромосом в одну гамету могут попасть обе гомологичные хромосомы из пары. 3. В этом случае гамета будет нести по паре аллелей всех генов, которые содержатся в данной паре хромосом. Аллельные гены, их взаимодействие. Множественные аллели. III. МНОЖЕСТВЕННЫЕ АЛЛЕЛИ. Большинство генов существует в виде двух вариантов аллелей. Но некоторые гены существуют в виде большего числа аллелей. Тогда в популяции имеется не два аллельных гена, а больше три, четыре до нескольких десятков. Такое явление называется множественным аллелизмом. Обозначаются множественные аллели одной буквой с указанием номера: А, а1, а2, а3, а4 Возникают они в результате многократных генных мутаций одного локуса гена. Чем больше аллельных генов, тем больше их комбинаций попарно. Они дают намного больше генотипов (Аа1, Аа2, Аа3, а1 а2, а1 а3, а2 а3, а3 а3), чем двухаллельные гены, дающие только три генотипа (АА, Аа, аа). У одной диплоидной особи может быть только два аллеля из серии множественных аллелей. Характер взаимодействия в серии множественных аллелей может быть по типу полного или неполного доминирования одного аллеля над другим, или кодоминирования. По типу множественных аллелей у человека наследуются: группы крови по системе АВО, цвет глаз, варианты серповидноклеточной анемии: НbA, HbS, HdC. Например, наследование групп крови по антигенам АВ. I – ген определяющий развитие групп крови по системе АВО. Множественные аллели групп крови по системе АВО обозначаются: IO, IA, IB. А и В – агглютиногены – сложные соединения, за синтез которых отвечает ген «I». По наличию и сочетанию агглютиногенов в крови различают четыре группы крови по системе АВО.
YI. ПЛЕЙОТРОПИЯ – это генетическое явление, обеспечивающее развитие нескольких признаков, контролируемых одним геном, проявляется множество фенотипических эффектов одного гена. Различают: а) прямую или истинную или первичную плейотропию б) зависимую или относительную или вторичную плейотропию a) При прямой плейотропии все разнообразные эффекты возникают в различных тканях или органах под действием одного гена в разных местах организма. Изменённый белок взаимодействует с клеточными структурами (мембраной, цитоплазмой) в клетках нескольких тканей и органов и изменяет их свойства. Например, синдром Марфана (арахнодактилия – «паучьи пальцы») у человека, развивается под действием мутантного доминантного гена - А, действие которого проявляется в нарушении развития соединительной ткани, что приводит к аномалии развития опорно-двигательного аппарата – длинные и тонкие пальцы, подвывиха хрусталика и развитие близорукости, аневризм (расширение) аорты уменьшает продолжительность жизни. Гомозиготы АА – нежизнеспособны, а у всех гетерозигот проявляются признака синдрома. б) Относительная (вторичная) имеется одно первичное действие мутантного гена, например, в случае серповидноклеточной анемии – дефектный гемоглобин и эритроциты принимают серповидную форму. Вследствие этого развивается ступенчатый процесс вторичных патологических проявлений - это дефекты, возникающие в различных органах: слипание и разрушение эритроцитов, анемия, умственная и физическая отсталость, увеличение и фиброз селезёнки, поражения кожи, почек, сердца, мозга. Поэтому гомозиготы (SS) по этому заболеванию погибают до полового созревания (в детском возрасте). Схема действия плейотропных генов Рассмотрим взаимодействия неаллельных генов расположенных в разных негомологичных хромосомах. Г. Менделю. Так, при взаимодействии двух генов для F2 используется общая формула расщепления по фенотипу 9/16 А-В, 3/16 А-вв, 3/16 ааВ-, 1/16 аавв. Но характерной особенностью взаимодействия неаллельных генов является развитие в потомстве новых вариаций признака, поэтому очень часто основные менделеевские расщепления модифицируются, т.к. некоторые генотипические классы по фенотипу не отличаются друг от друга. Таким образом, различные типы взаимодействия генов получаются из расщепления 9:3:3:1 путём объединения его элементов. Например: 9:3:4 = 9: 3: (3+1), 9:7 = 9: (3:3:1), 12:3:1 = (9+3):3:1. Необходимо лишь помнить о характере взаимодействия и правильно определять фенотип потомства и родителей. Расщепление может быть: ü при комплементарном взаимодействии - 9:7, 9:6:1, 9:3:4, ü при эпистазе - 13:3, 12:3:1, ü при полимерном (аккумулятивное действие) - 1:4:6:4:1 или 15:1 (когда гены не усиливают друг друга). Рассмотрим взаимодействие неаллельных генов (комплементарность, эпистаз, полимерия) Комплементарность – это взаимодействие неаллельных генов, когда два доминантных гена А и В встретившись в зиготе при оплодотворении определяют развитие нового признака. При этом каждый из комплементарных генов в отдельности не обладает способностью обеспечить развитие данного признака. Наследование, сцепленное с полом Исследования показали, что наследование некоторых признаков происходит не в соответствии с менделевскими закономерностями, а обнаруживает отчетливую связь с полом. Если при обычном менделевском наследовании оба родителя в равной мере могут передать признак детям, то для признаков, сцепленных с полом, результаты наследования будут различными в зависимости от того, кто обладал данным признаком – мать или отец. По мере изучения наследования признаков у различных организмов было замечено, что не все гены подчиняются менделевскому правилу свободного комбинирования. Некоторые гены обнаруживают отчетливую тенденцию к совместному или сцепленному наследованию. Это явление было впервые замечено в 1906 г У. Бэтсеном и Р. Пеннетом они пришли к заключению, что действие третьего з-на Менделя является ограниченным в связи с существованием явления притяжения и отталкивания генов. Черн. корот. сер. норм. Р ♀ ввvv Х ♂ F1ВвVv
Ġ вv ВV вv 50% 50% F2 Вв Vv вв vv Сер. норм. черн. корот. 1: 1 У гибридной самки дрозофилы ♀ BbVv, напротив, благодаря наличию кроссинговера, обнаруживается неполное сцепление генов. При этом у нее образуются четыре типа гамет со всеми возможными комбинациями генов, но количество их неравное. Большую часть среди них составляют некроссоверные гаметы (83%), несущие хромосомы с исходным родительским положением генов, и меньшую часть - кроссоверные гаметы(17%), содержащие рекомбинантные хромосомы с новыми комбинациями генов.
Р ♀ Вв Vv х ♂ вв vv
Ġ ВV вv в V Вv вv 41,5% 41,5% 8,5% 8,5% некроссоверные кроссоверные гаметы гаметы
F2 Вв Vv вв vv Ввv v ввVv сер. норм. черн. корот. сер. корот. черн. норм. 41,5% 41,5% 8,5% 8,5%
некроссоверные кроссоверные
Количество кроссоверных гамет и, соответственно, кроссоверных форм в потомстве зависит от частоты кроссинговера, а она в свою очередь, - от расстояния между генами. Чем больше расстояние между генами, тем выше вероятность кроссинговера между ними и меньше сила сцепления, и наоборот. Закон Моргана: «Г ены, расположенные в одной хромосоме, наследуется, сцеплено, причём сила сцепления между генами обратно пропорциональна расстоянию между ними».
Открытие кроссинговера позволило разработать принцип построения генетических карт хромосом. Явление сцепления генов было использовано для выяснения локализации генов, расположенных в одной хромосоме. На основании того, что частота кроссинговера между отдельными генами в группе сцепления постоянна. Морган предложил оценивать расстояние между генами в хромосоме в относительных единицах – процентах (%) кроссинговера (по предложению советского генетика А.С.Серебровского единица, равная 1% кроссинговера, была названа морганидой). Определения между сцепленными генами проводиться путем подсчета суммарного процента кроссоверных особей в потомстве анализирующего с крещивания. Поскольку в опыте Моргана количество рекомбинантных форм по окраске тела и длине крыльев у дрозофилы составило 17% кроссинговера (или 17 морганидам). Определение аналогичным путем расстояния между другими генами в группах сцепления обнаружило линейный порядок расположения генов в хромосоме. Это позволило установить взаимное расположение генов в группах сцепления и составить генетические карты хромосом. Генетическая карта хромосом является графическим отображением линейной последовательности расположения генов в группе сцепления А в с д Е р О К Она не дает точного представления об истинном физическом расстоянии между генами в хромосоме, так как частота кроссинговера неодинакова по длине хромосомы (например, в околоцентромерных районах кроссинговер происходит очень редко). Чем меньше расстояние между генами в хромосоме, тем точнее процент кроссинговера отражает фактическое расстояние между ними. При больших расстояниях между генами множественный кроссинговер искажает истинное расстояние между ними. В связи с этим при составлении генетических карт хромосом расстояния между далеко отстоящими генами определяются не по частоте кроссинговера между ними, а путем сложения расстояний (в % кроссинговера) между многими промежуточными генами. Поэтому общая длина хромосомы на генетической карте может превышать 100%. В настоящее время генетические карты хромосом построены для многих биологических объектов: дрозофилы, кукурузы, томатов, мышей, в том числе и для человека. У человека анализ сцепления генов классическими методами невозможен. Для составления карт хромосом используются: генеалогический метод, основанный на анализе родословных, методы молекулярной биологии, генной инженерии, дифференциальной окраски хромосом, культивирования гибридных соматических клеток человек-мышь. ГЕНЕТИКА ЧЕЛОВЕКА Генетика человека сформировалась с учетом особенностей, создающих трудности при изучении наследственности и изменчивости человека. ГЕНЕАЛОГИЧЕСКИЙ МЕТОД Основой метода является составление родословной и её последующий анализ. Задачами этого метода является установление наследственного характера признака и определение типа наследования. Даёт возможность изучать явление сцепленного наследования и взаимодействия генов, определить пенетрантность аллелей. Он лежит в основе медико-генетического консультирования. Составление родословной – это сбор сведений о семье. Начинается с человека, обратившегося в врачу – генетику. Человек с которого начинают составлять родословную называется п ро б а н д о м. При составлении родословной используют стандартные символы. Поколения в родословной обозначают римскими цифрами, которые ставят слева от родословной. Арабскими цифрами нумеруют потомство одного поколения слева направо. Братья и сестры располагаются в порядке рождения. СИМВОЛЫ РОДОСЛОВНОЙ.
Определение пенетрантности. Пенетрантность гена, по формуле K = 5. Определение вероятности рождения ребенка в семье с наследственной патологией (на основе решения генетической задачи) 6. Определение взаимодействия аллельных и неаллельных генов и их тип (полное доминирование, неполное доминирование, кодоминирование, эпистаз, комплементарность)
ДЕРМАТОГЛИФИЧЕСКИЙ МЕТОД Дерматоглифика - наука, изучающая узоры на коже. Наука о папиллярных линиях, контролируемых генотипом, узоры индивидуальны и сохраняются в течение всей жизни, их практически невозможно удалить. Поэтому их изучение используют для определения зиготности близнецов, идентификации личности, для установления отцовства, в криминалистике, для диагностики хромосомных болезней. Классификация: - Дактилоскопия – изучение узоров на подушечках пальцев - Пальмоскопия – изучение узоров на ладонях - Плантоскопия – изучение узоров на подошве стопы
ДАКТИЛОСКОПИЧЕСКИЕ показатели: 1. Тип узора (качественный показатель) 2. Дельтовый индекс (количественный показатель) 3. Гребневой счёт (количественный показатель) 1. Типы узоров:
А – дуга полузамкнутый простой рисунок L -полузамкнутый узор, различают по направлению открытого конца – ульнарный (LU – петля ульнарная, рисунок открывается в сторону мизинца) и радиальный (LR – петля радиальная, рисунок открывается в сторону большого пальца) W – концентрический узор Узорная формула: D (правая рука) - I II III IV V W LR A 2L LU S (левая рука) - I II III IV V W LU LU W A Дельтовый индекс Dl – дельта (треугольник) образуется в результате схождения папиллярных линий, идущих из 3-ех направлений и нигде не пресекающихся. Дельтовый индекс равен сумме дельт на 10 пальцах и колеблется от 0 до 20 дельт A – дуга не имеет трирадиуса Dl = 0 LU, LR – петля имеет один трирадиус Dl = 1 2L – двойная петля имеет два трирадиуса Dl = 2 W – завиток имеет два трирадиуса Dl = 2
3. Гребневой счёт (Q) – это количество папиллярных линий, пересекающих прямую проведенную из центра рисунка в центр радиуса.
В норме гребневой счёт равен Q10 = 127±50 ♀ Q10 = 145±50 ♂
1. Ладонные подушечки – это шесть возвышений окружающих центральную ямку на ладони: а. Тенар б. Гипотенар в. Ладонные межпальцевые подушечки (4 штуки)
.
2. Ладонные складки (В норме – 5, реже 4)
При соединении дистальной и проксимальной складок образуется ЧПЛ четырёхпальцевая складка или борозда ЧПБ. Количество ладонных складок при этом уменьшается с 5 до 4. Образование ЧПЛ происходит в норме 5-6% случаях. 100% характерно для хромосомных синдромов. Ладонные поля
4. Ладонные пальцевые трирадиусы (треугольники), лежат у основания I I – Y - пальцев
II III IY Y
a b c d дельта, трирадиусы обозначаются прописными буквами лат. алфавита (a, b, c, d)
Первые a, b, в норме всегда присутствуют, c и d могут отсутствовать. Также могут появляться дополнительные межпальцевые трирадиусы a1, b1, c1 d1
5. Главные ладонные линии - обозначаются заглавными буквами латинского алфавита (А, B,С, D)
Они начинаются от ладонных трирадиусов a b c d и заканчиваются в определенных полях. Формула для главных ладонных линий: A B C D 5’ 7 9 11
6. Индекс Камминса – сумма полей выхода главных ладонных линий A и D по индексу полей Камминса ИК= А + D
A B C D ® главные ладонные линии 5 6 → номер поля по Камминсу ИК = 5 + 6 = 11 7. Угол atd - главный осевой ладонный трирадиус.
Трирадиус t находится в проксимальной части ладони в близи браслетной складки. Угол atd, образуется при пересечении линий проведённых от трирадиусов a и d к трирадиусу t. В норме угол atd равен 57°, при болезни Дауна — 81°, при трисомии по другим аутосомам — 108°.
t – трирадиус, который расположен к проксимальному краю ладони.
t
Аутосомные анеуплоидии 1) 47,+21,xx – с. Дауна (ЧПЛ,
2) 47,+18,ху(хх) – с. Патау (LR>LU,
3) 47,+18, хх(ху) с. Эдвардса (подушечки слабо выражены, только дуги А, ЧПЛ, Гоносомные: С. Тернера-Шерешевского ♀ 45, хо (W>L, возрастает Q и Dl, atd смещается дистально и ульнарно С. Клайнфельтера ♂ 47,хху (А больше, Dl и Q снижается,
Цитогенетический метод. Цитогенетический метод основан на микроскопическом исследовании хромосом – · кариотипа (индивидуального набора хромосом в норме, а также при геномных и хромосомных мутациях) · полового хроматина Позволяет выявлять
а) полиплоидии, б) анеуплоидии – трисомии 2п + 1 (с. Дауна, Эдварса, Патау, Клайнфельтера), - моносомии 2п – 1 (Тернера - Шерешевского), - нулисомии 2п – 2 в) мозаицизм 46/45 хромосом в группах клеток
Этапы цитогенетического метода:
Основные сведения о морфологии хромосом человека получены при изучении их в метафазах митоза прометафазе-метафазе мейоза. При этом важно, чтобы количество делящихся клеток, было достаточно велико. Окрашивание. Наиболее простой способ окрашивания – простая рутинная сплошная по всей длине хромосомы основным (щелочным) красителем Гимза или 2% - ным ацетоорсеином или ацеткармином. Эти красители окрашивают хромосомы целиком, равномерно и интенсивно. Для выявления численных аномалий хромосом этот метод вполне достаточен. Для получения более детальной картины структуры хромосом или их сегментов используют различные способы дифференциального окрашивания. Методы дифференциальной окраски хромосом основаны на действии солевых растворов с определённым Ph, температурным режимом, обработкой ферментами протеазами. Этими методами установлена четкая структурная разнородность хромосом по длине на красящиеся (тёмные - гетерохроматин) и некрасящиеся (светлые - эухроматин) полосы. Дифференциальное окрашивание приводит к появлению линейного рисунка по длине хромосомы. Окрашивание может охватывать отдельные районы. Рисунок этих полос специфичен, индивидуален для каждой пары хромосом. Рисунок сегментации зависит от особенностей неоднородности целостного комплекса ДНК – белок в разных участках по длине хромосом. Различные типы сегментов обозначают по методам, с помощью которых они выявляются наиболее отчетливо. G R Классификация хромосом Группа А (1 – 3) – 6 хромосом, три наиболее крупных хромосом: первая и третья пары метацентрические, а вторая умеренно субметацентрическая Группа В (4 - 5) – 4 хромосомы, две пары длинных больших субметацентриков, неразличимых по размеру и морфологическим признакам. Группа С (6 - 12) – 14 хромосом, семь пар аутосом средней величины с субмедианно расположенной центромерой. К этой группе относится половая Х-хромосома, она сходна с самыми крупными хромосомами из группы С – 6 и 7. Группа Д (13 - 15) – 6 хромосом, три пары больших (средней величины) акроцентриков не различимых между собой. Группа Е (16 - 18) – 6 хромосом, три пары малых субметацентрических хромосом 16 – хромосома имеет почти медианную локализацию центромеры, а 18 –пара хромосом более акроцентрична, т.е отлтчается меньшей длиной коротких плеч Группа F (19 - 20) – 4 хромосомы, две пары маленьких метацентриков Группа G (21 - 22) – 4 аутосомы, две пары мелких акроцентриков. Эти хромосомы неразличимы. К этой группе относится У-хромосома (мелкий акроцентрик) Половые хромосомы Х и У распределяются в конце идеограммы. Символика хромосом Кариотип человека в норме и при хромосомных заболеваниях требуют унифицированной символики хромосом. В настоящее время исследователи всего мира – клинические генетики, невропатологи, педиатры, психологи, психиатры используют «Международную систему для цитогенетической номенклатуры хромосом человека». Согласно последней номенклатуре такие морфологические признаки хромосом, как теломеры, центромеры, специфические полосы по длине хромосомы, используются в качестве сравнительных знаков. В хромосоме выделяют - короткое – p и длинное - q плечи хромосом. В каждом плече выделяют районы, полосы и сегменты, которые пронумерованы от центромеры к теломеры, например, запись – 6 q 2,3 указывает на то, что это хромосома 6, длинное плечо q, район 2, сегмент 3. Районы и сегменты на хромосомах проявляются после дифференциального окрашивания хромосом различными методами Q, G, R, C, T. В номенклатуре существует следующие обозначения для описания нормальных кариотипов 46, ХХ девочка и 46, ХУ мальчик. В начале кариотипа записывается общее число хромосом, включая половые хромосомы. Затем записываем половые хромосомы. Численные аномалии обозначаются изменением числа хромосом в кариотипе и указанием (+) или (–) той или иной присутствующей или отсутствующей хромосомы. Исключением являются половые хромосомы, при количественных аномалиях которых (+) или (–) никогда не ставятся. Причём отсутствие одного из гомологов хромосом или его части (делеция) в кариотипе, несмотря на тип мутации (численной или структурной), носит название моносомии – полной или частичной, а присутствие – трисомии полной или частичной. Полиплоидии о тражаются только числом хромосом (69). Мозаичные формы Эта группа заболеваний, вызванная соматическими мутациями на первых стадиях онтогенеза – дробления. Дети – мозаики могут появляться у здоровых родителей. При мозаицизме mos обозначают кариотип каждoго аномального и нормального клона клеток, после чего записывают в скобках [ ]. Число проанализированных клеток, кариотипы разных клонов обозначают косой чертой /. Нормальный клон всегда обозначается последним Мозаичный синдром Дауна mos 47,ХХ, +21[60]/46,ХХ[40], запись кариотипа говорит о том, что у девочки с мозаичной формой с. Дауна при анализе 100 клеток обнаружено 40 клеток с нормальным кариотипом и 60 клеток с простой трисомией по 21 хромосоме. Мозаичный синдром Тернера 45,ХО, [80]/ 46,ХХ [20] или 45,ХО[30]/ 47,ХХХ[70] ПОЛОВОЙ ХРОМАТИН В 1949 г. Барр и Бертрам в ядрах нейронов самок кошек обнаружили интенсивно, окрашенную глыбку хроматина. Она имеет треугольную форму и прилежит к внутренней мембране ядра. Далее было установлено, что только одна из двух Х-хромосом в соматических клетках женских особей функционально активна, другая конденсируется (факультативные гетерохроматин), и в интерфазном ядре образует Х-хроматин, или тельце Барра. Этот процесс имеет случайный характер в эмбриональном периоде. В норме у женщин (46, ХХ)– одно тельце Барра, у мужчин (46, ХУ) оно отсутствует. Изменение числа Х-хромосом ведёт к изменению числа телец Барра. Методы моделирования Разработаны два метода: биологическое и математическое моделирование. С помощью этих методов решаются разные задачи, имеющее значение как для разработки теоретических основ генетики человека, так и для практического медико-биологического консультирования. Биологическое моделирование – это использование животных, имеющих наследованные аномалии, соответствующие аномалиям человека. Используется в генетике для: 1. изучения патогенеза наследственных заболеваний. 2. разработки методов их лечения. Математическое моделирование – основано на использовании компьютерных технологий. С помощью этого метода изучаются: 1. Распространение мутаций в популяциях при разных условиях действие отбора; 2. Влияние экспериментальных эволюционных факторов в разных сочетаниях на генофонд популяций при распространении наследственной патологии.
Близнецовый метод Это метод изучения генетических закономерностей на близнецах. Впервые он был предложен Ф. Гальтоном в 1875 г. Близнецовый метод дает возможность определить вклад генетических (наследственных) и средовых факторов (климат, питание, обучение, воспитание и др.) в развитии конкретных признаков или заболеваний у человека. При использовании близнецового метода проводится сравнение: 1) монозиготных (однояйцевых) близнецов — МБ с дизиготными (разнояйцевыми) близнецами — ДБ; 2) партнеров в монозиготных парах между собой; 3) данных анализа близнецовой выборки с обшей популяцией. Монозиготные близнецы образуются из одной зиготы, разделившейся на стадии дробления на две (или более) части. С генетической точки зрения они идентичны, т.е. обладают одинаковыми генотипами. Монозиготные близнецы всегда одного пола. Имеют одну плаценту. Особую группу среди МБ составляют необычные типы близнецов: двухголовые (как правило, нежизнеспособные), каспофаги ("сиамские близнецы"). Наиболее известный случай — родившиеся в 1811 г. в Сиаме (ныне Таиланд) сиамские близнецы — Чапг и Эиг. Они прожили 63 года, были женаты на сестрах-близнецах; Чанг произвел на свет 10, а Энг - 12 детей. Когда от бронхита умер Чанг, спустя 2 часа умер и Энг. Их связывала тканевая перемычка шириной около 10 см от грудины до пупка. Позднее было установлено, что соединявшая их перемычка содержала печеночную ткань, связывающую две печени. Любая хирургическая попытка разделить братьев вряд ли в то время была бы успешной. В настоящее время разъединяют и более сложные связи между близнецами. Дизиготные близнецы развиваются в том случае, если одновременно две яйцеклетки оплодотворены двумя сперматозоидами. Естественно, дизиготные близнецы имеют различные генотипы. Они сходны между собой не более, чем братья и сестры, т.к. имеют около 50 % идентичных генов. Общая частота рождения близнецов составляет примерно 1 %, из них около 1/3 приходится на монозиготных близнецов. Известно, что число рождений монозиготных близнецов сходно в разных популяциях, в то время как для дизиготных эта цифра существенно различается. Например, в США дизиготные близнецы рождаются чаще среди негров, чем белых. В Европе частота появления дизиготных близнецов составляет 8 на 1000 рождений. Однако в отдельных популяциях их бывает больше. Самая низкая частота рождения близнецов присуща монголоидным популяциям, особенно в Японии. Отмечается, что частота врожденных уродств у близнецов, как правило, выше, чем у одиночно рожденных. Полагают, что многоплодие генетически обусловлено. Однако это справедливо лишь для дизиготных близнецов. Факторы, влияющие на частоту рождения близнецов, в настоящее время мало изучены. Есть данные, показывающие, что вероятность рождения дизиготных близнецов повышается с увеличением возраста матери, а так же порядкового номера рождения. Влияние возраста матери объясняется, вероятно, повышением уровня гонадотропина, что приводит к учащению полиовуляцни. Имеются также да
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-25; просмотров: 260; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 13.59.67.189 (0.012 с.) |







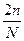 х 100%, где К – пенетрантность, n – количество больных потомков, N – общее количество потомков.
х 100%, где К – пенетрантность, n – количество больных потомков, N – общее количество потомков.



 ПАЛЬМОСКОПИЯ:
ПАЛЬМОСКОПИЯ:






 atd = 57º (норма)
atd = 57º (норма) 81º (с. Дауна)
81º (с. Дауна)
 a d
a d



