Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Последовательность событий инициации трансляцииСодержание книги
Поиск на нашем сайте
Этап 1. Диссоциация нетранслирующих (терминирующих) 70S рибосом на 30S и 50S субъединицы предшествует инициации трансляции во всех случаях. При физиологических условиях нетранслирующие рибосомы, по всей вероятности, находятся в состоянии обратимого равновесия со своими отдельными субъединицами. IF1 может ускорить процесс диссоциации/ассоциации, в то время как IF3 связывается с 30S субъединицами и нарушает состояние равновесия: 70S ⇔ 30S + 50S … + IF3 -> 30S:IF3 Этап 2. 30S рибосомные субъединицы с IF3 или со всеми тремя факторами инициации («нативные» 30S частицы) могут связываться с участком инициации, или участком связывания с рибосомой (RBS), на мРНК. Этот участок может быть расположен вблизи от 5'-конца или далеко от него; в случае полицистронной мРНК возможно существование нескольких таких участков на протяжении цепи. Важно то, чтобы этот участок мРНК был доступен для взаимодействия с рибосомной частицей и содержал полипуриновую последовательность Шайна-Дальгарно (SD) и на должном расстоянии за ней - триплет инициации (предпочтительно AUG). В отсутствие факторов инициации 30S рибосомная субъединица также способна распознавать участок инициации (RBS) на мРНК, но факторы существенно усиливают взаимодействие и делают его гораздо более избирательным: 30S:(IF3:IF1:IF2) + mRNA -> mRNA:30S:(IF3:IF1:IF2). мРНК, связанная в этом комплексе, удерживается в основном взаимодействием SD:ASD и еще не установлена полностью и должным образом в мРНК-связывающей щели 30S субъединицы («состояние ожидания» - “stand-by state”). Этап 3. На этом этапе, в соответствии с классическим сценарием, описанным выше, F-Met-тРНК связывается с комплексом мРНК:30S. Это связывание является ГТФ-зависимым и опосредовано IF2. Если IF2 уже находится на 30S частице, он активируется при помощи ГТФ и, таким образом, приобретает сродство к F-Met-тРНК, и F-Met-тРНК присоединяется к комплексу: mRNA:30S:IF3:IF1:IF2 + GTP + F-Met-tRNA ® ® mRNA:30S:IF3:IF1:IF2:GTP:F-Met-tRNA. Имеются веские основания считать, что кодон-антикодоновое взаимодействие инициаторного триплета мРНК и связанной F-Met-тРНКfMet окончательно устанавливает мРНК в мРНК-связывающем участке 30S рибосомной субъединицы (см. раздел 8.2). Если свободный IF2 с ГТФ случайно сталкивается с F-Met-тРНК в растворе, они сначала образуют тройственный комплекс IF2:GTP:F-Met-tRNA, который затем связывается с комплексом mRNA:30S:
mRNA:30S:IF3:IF1 + F-Met-tRNA:IF2:GTP ® ® mRNA:30S:IF3:IF1:IF2:GTP:F-Met-tRNA. Также может обсуждаться и альтернативный путь, когда «нативная» 30S частица с факторами инициации сначала связывает F-Met-тРНК, а затем сформированный комплекс взаимодействует с участком связывания с рибосомой на мРНК: 30S:IF3:IF1:IF2:GTP + F-Met-tRNA ® 30S:IF3:IF1:IF2:GTP:F-Met-tRNA; 30S:IF3:IF1:IF2:GTP:F-Met-tRNA + mRNA ® ® mRNA:30S:IF3:IF1:IF2:GTP:F-Met-tRNA. В любом случае, именно антикодон F-Met-тРНК, связанной на 30S субъединице, фиксируется на инициаторном кодоне, расположенном за полипуриновой последовательностью, и тем самым устанавливает рибосомную частицу точно на начало кодирующей последовательности. Этап 4. Теперь инициаторный 30S комплекс готов объединиться с 50S рибосомной субъединицей. Факторы инициации, естественно, должны освобождаться из комплекса как-то сопряженно с ассоциацией субъединиц. Фактор-связывающий участок 50S субъединицы взаимодействует с IF2 и индуцирует ГТФазную активность этого фактора. ГТФ гидролизуется, что приводит к потере белком IF2 сродства к F-Met-тРНК и к рибосоме. Меньше известно о моменте и механизме освобождения двух других факторов инициации. Так или иначе, 70S частица с инициаторной F-Met-тРНК в Р участке формируется непосредственно на инициаторном кодоне мРНК: mRNA:30S:IF3:IF1:IF2:GTP:F-Met-tRNA + 50S -> -> [mRNA:30S:IF3:IF1:IF2:GTP:F-Met-tRNA:50S] -> -> mRNA:70S:F-Met-tRNA + IF3 + IF1 + IF2 + GDP + Pi. Этап 5. Инициаторный 70S комплекс, сформированный в предыдущей реакции, содержит вакантный А участок с установленным в нем кодоном, следующим непосредственно за инициаторным кодоном. Р участок занят инициаторной F-Met-tRNA - аналогом пептидил-тРНК. Таким образом, 70S комплекс компетентен к принятию первой элонгаторной аминоацил-тРНК в свой вакантный А участок и осуществляет формирование первой пептидной связи между двумя субстратами: инициаторной F-Met-тРНК и первой элонгаторной аминоацил-тРНК: mRNA:70S:F-Met-tRNAf + Aa-tRNAe:EF-Tu:GTP -> -> mRNA:70S:F-Met-tRNAf:Aa-tRNAe:EF-Tu:GTP -> -> mRNA:70S:F-Met-tRNAf:Aa-tRNAe + EF-Tu:GDP + Pi -> -> mRNA:70S:F-Met-Aa-tRNAe + tRNAfMet + EF-Tu:GDP + Pi.
На этом инициация завершается, и начинается элонгация.
30. Конститутивный контроль трансляции на стадии инициации. (Бейлин) Скорость или частота инициации трансляции рибосомами может сильно различаться для разных мРНК. У прокариотических организмов это определяется тем, что инициирующие или рибосомосвязывающие участки разных мРНК имеют разное сродство к рибосомам и, таким образом, с разной эффективностью связывают рибосомные частицы. На основании разницы в эффективности инициации можно говорить о <сильных> и <слабых> мРНК. На сильных мРНК инициация происходит часто, на них нанизывается много рибосом (образуются плотные полирибосомы) и соответственно продуцируется много молекул белка. Редкая инициация трансляции на слабых мРНК дает в результате редкую посадку рибосом на эти мРНК и, следовательно, низкую белковую продукцию.
Известной усиливающей последовательностью, расположенной по направлению к 5’-концу от последовательности Шайна-Дальгарно, является так называемый «эпсилон-мотив» UUUAACUUUA высокоэкспрессируемых поздних мРНК бактериофага Т7 и некоторых других фагов, а также эпсилон-подобные последовательности некоторых бактериальных мРНК - например, UUUUAACU и UAAUUUAC в высокоэкспрессируемом цистроне atpE полицистронной atp-мРНК у E. сoli. Похожая ситуация наблюдается и в эукариотических клетках, но там дискриминация мРНК обусловлена скорее разным сродством факторов инициации, а не самих рибосом к разным 5'-проксимальным инициаторным структурам мРНК. Так как факторы инициации в любом случае локализуются на инициирующих малых рибосомных субчастицах, то они и определяют разную эффективность посадки рибосом на разные мРНК и, таким образом, дискриминируют их на сильные и слабые. Различная сила мРНК в значительной мере определяет соотношение продукции различных белков в клетке. ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ «СИЛУ» мРНК (1) Инициаторный кодон: AUG > GUG > UUG > AUA, etc. (2) SD: сила спаривания с рРНК. (3) Расстояние между AUG и SD (4 – 12 нуклеотидов – оптимально). (4) «Энхансерные» последовательности «вверх» от SD. (5) Вовлеченность AUG и SD во вторичную и /или третичную структуру.
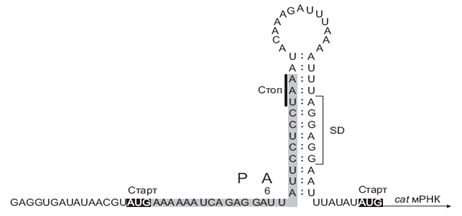
31. Индуцированная инициация трансляции. Реинициация. (Бейлин) Схема механизма индукции трансляции мРНК, кодирующей бактериальную хлорамфеникол-ацетилтрансферазу (cat-мРНК), хлорамфениколом
Последовательность Шайна-Дальгарно (SD) этой мРНК блокирована стабильной вторичной структурой, образованной спариванием с концом предшествующей короткой открытой рамки считывания (sORF), так что независимая инициация трансляции мРНК невозможна. В отсутствие хлорамфеникола рибосомы транслируют предшествующую ORF вплоть до терминирующего кодона UAA и расплетают межцистроновую шпильку, но из-за чересчур короткого расстояния между UAA и SD транслирующие и терминирующие рибосомы мешают инициирующим субъединицам взаимодействовать с SD. В присутствии хлорамфеникола трансляция sORF ингибируется, и рибосомы останавливаются в середине sORF (по неизвестным причинам, предпочтительно на шестом кодоне); в этой ситуации межцистроновая шпилька оказывается уже расплетенной и рибосомо-связывающий участок cat-мРНК открытым для инициации.
|
||||||
|
|
Последнее изменение этой страницы: 2016-08-16; просмотров: 477; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.15.231.199 (0.011 с.) |