Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Нарушения со стороны вестибулярной системы.Содержание книги Поиск на нашем сайте
Сильные раздражения вестибулярной системы часто связаны с возникновением неприятных ощущений, головокружением, рвотой, усиленным потоотделением, тахикардией и т. д. В таких случаях говорят о кинетозе (укачивание- «морская болезнь») Укачивание, вероятно, является следствием комплекса стимулов, необычных для организма (например, в море). Действие кориолисова ускорения особенно сильно проявляется в условиях расхождения между зрительным восприятием и сигналами вестибулярного аппарата. У новорожденных, а также больных, не имеющих лабиринта, укачивания не наблюдается. Острое одностороннее нарушение функции лабиринта вызывает тошноту, рвоту, потливость, головокружение и иногда нистагм, направленный в здоровую сторону. У больных наблюдается тенденция к падению на ту сторону тела, где нарушена лабиринтная функция. Очень часто, однако, клиническая картина осложнена неопределенностью направления головокружения, нистагма и падений. При некоторых заболеваниях, таких, как синдром Меньера, иногда возникает избыточное давление эндолимфы с одной из сторон, при этом первым последствием раздражения рецепторов оказываются симптомы, противоположные по направлению вызываемым описанной выше деструкцией рецепторов. В противоположность грубым проявлениям острых вестибулярных заболеваний хроническое выпадение функции лабиринта компенсируется сравнительно хорошо. Деятельность центральной вестибулярной системы может перестраиваться таким образом, чтобы ослабить реакцию организма на патологическую афферентацию. Такая регуляция особенно точна, когда другие сенсорные, например зрительные или тактильные, каналы обеспечивают корректирующие афферентные импульсы. Поэтому патологические проявления, связанные с хроническими вестибулярными расстройствами, более выражены, например, в темноте. Острое двустороннее нарушение у человека встречается редко. В опытах на животных возникающие нарушения выражены намного сильнее, чем при одностороннем разрушении; если предотвратить афферентацию в вестибулярные ядра с обеих сторон, наблюдаются симметричные симптомы. Проприоцептивный АНАЛИЗАТОР Двигательный (проприоцептивный) анализатор обеспечивает формирование так называемого «мышечного чувства» при изменении напряжения мышц, их оболочек, суставных сумок, связок, сухожилий. В мышечном чувстве можно выделить три составляющих: чувство положения, когда человек может определить положение своих конечностей и их частей относительно друг друга; чувство движения, когда, изменяя угол сгибания в суставе, человек осознает скорость и направление движения; чувство силы, когда человек может оценить мышечную силу, нужную для движения или удерживания суставов в определенном положении при подъеме или перемещении груза. Наряду с кожным, зрительным, вестибулярным двигательный анализатор оценивает положение тела в пространстве, позу, участвует в координации мышечной деятельности. Периферический отдел представлен проприорецепторами, расположенными в мышцах, связках, сухожилиях, суставных сумках, фасциях. К ним относятся мышечные веретена, тельца Гольджи, тельца Фатера—Пачини, свободные нервные окончания. Мышечное веретено представляет собой скопление тонких коротких поперечнополосатых мышечных волокон, которые окружены соединительнотканной капсулой. Эти волокна получили название интрафузальных в отличие от обычных мышечных волокон, которые составляют основную массу мышц и называются экстрафузальными, или рабочими, волокнами. Интрафузальные волокна расположены параллельно экстрафузальным, поэтому возбуждаются при расслаблении (удлинении) скелетной мышцы. Рассмотрим подробно миотатический рефлекс. В мышце есть три основных типа проприорецепторов: 1) первичные окончания веретён; 2) вторичные окончания веретён; Рецепторы Гольджи.
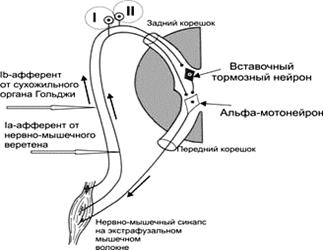
Мышечное веретено длиной 3 мм шириной 0,3 мм состоит из интрафузальных мышечных волокон, которые окружены толстой соединительнотканной капсулой, внутри которого находится жидкость, подобная лимфе. Два толстых волокна имеют по экватору скопление ядер (ядерная сумка), и волокна называются ядерно-сумчатыми. Четыре тонких волокна имеют по экватору ядра расположенные в один ряд (ядерная цепочка), а волокна получили название ядерно-цепочечных. В разных скелетных мышцах число веретен на 1 г ткани варьирует от нескольких единиц до сотни. На интрафузальных волокнах спирально расположены чувствительные окончания афферентных волокон группы Iа — Эти волокна называются первичными афферентами. Первичные окончания (около 15мкм) -миелинизированное чувствительное нервное волокно Ia (скорость проведения 70–120 м/с) формирует первичное, или аннулоспиральное окончание, (они представляют из себя рецепторную спираль). Эти окончания возбуждаются при расслаблении или удлинении мышцы, вызванной растяжением, и реагируют на скорость удлинения экстрафузальных волокон. При механической деформации аннуло-спиральных окончаний (периферическое воздействие) рождается рецепторный потенциал, который далее преобразуется в потенциал действия и распространяется по толстым афферентным волокнам первой группы ( Iа ) к спинальным ганглиям. Далее волокна от спинального ганглия направляются к а-фазическим нейронам и от них к белым мышцам (быстрым нейромоторным единицам). Импульсация, идущая от веретен по афферентным волокнам группы Iа в спинном мозге моносинаптически возбуждает мотонейроны своей мышцы и через тормозящий интернейронтормозит мотонейроны мышцы-антагониста ( реципрокное торможение). Аннуло-спиральные терминали могут также возбуждаться (в ответ на их деформацию) при сокращении интрафузальных мышечных волокон под влиянием нервных импульсов от γ-нейронов (центральное воздействие). К ядерно-цепочечным волокнам также подходят чувствительные окончания афферентных волокон группы II - (образовано 1–2 тонкими чувствительными нервными волокнами диаметром около 8мкм.) — так называемые вторичные окончания которые заканчиваются на них гроздьюбляшек - это колечки, розетки, утолщения терминалей. Они механически деформируются и возбуждаются под влиянием статической длины мышцы. Афферентные волокна группы II возбуждают мотонейроны мышц-сгибателей и тормозят мотонейроны мышц-разгибателей. Имеются, однако, данные, что афферентные волокна группы II, идущие от мышц-разгибателей, могут возбуждать мотонейроны своей мышцы. Потенциалы действия, пришедшие к атоническим нейронам по волокнам второй группы, далее направляются к красным мышцам (медленным нейромоторным единицам). Гамма-регуляция мышечного тонуса. Степень сокращения (напряжения) мышцы зависит от частоты импульсов, приходящих к ней от а-мотонейронов. Частота разрядов а-мотонейронов в свою очередь регулируется импульсами от проприорецепторов этой же мышцы. Веретена имеют и эфферентную иннервацию: интрафузальные мышечные волокна иннервируются аксонами, идущими к ним от γ -мотонейронов. Эти так называемые γ -эфферентные волокна подразделяют на динамические и статические. Разряды γ-мотонейронов повышают чувствительность мышечных веретён, причем динамические γ -эфференты преимущественно усиливают реакцию на скорость удлинения мышцы, а статические — на длину. Активация у-эфферентов и без растяжения мышцы сама по себе вызывает импульсацию афферентов веретен вследствие сокращения интрафузальных мышечных волокон. В результате получается, что при одинаковой длине мышцы увеличивается поток импульсов от рецепторов к а-мотонейронам, а от них - к мышце. Тем самым повышается мышечный тонус. Уровень возбуждения γ -системы тем выше, чем интенсивнее возбуждены а-мотонейроны данной мышцы, т. е. чем больше сила ее сокращения. Сами γ -мотонейроны контролируются ретикулярной формацией ствола мозга, мозжечком и корой. Благодаря сложному строению мышечные веретёна реагируют не только на периферические возбуждения (изменение длины мышцы), но и на изменения активных центральных механизмов - возбуждение γ -мотонейронов («γ -петля»). Таким образом, веретена можно рассматривать как непосредственный источник информации о длине мышцы и ее изменениях, если только мышца не возбуждена. При активном состоянии мышцы необходимо учитывать влияние у-системы. Во время активных движений γ -мотонейроны поддерживают импульсацию веретен укорачивающейся мышцы, что дает возможность рецепторам реагировать на неравномерности движения как увеличением, так и уменьшением частоты импульсации и участвовать таким образом в коррекции движений. Сухожильные рецепторы Гольджи (тельца Гольджи) находятся в сухожилиях. Это гроздьевидные чувствительные окончания, достигающие у человека 2—3 мм в длину и 1 — 1,5 мм в ширину. Тельца Гольджи, располагаясь в сухожилиях, включены относительно скелетной мышцы последовательно, поэтому они возбуждаются при ее сокращении вследствие натяжения сухожилия мышцы. Они имеют высокий порог возбуждения. Рецепторы Гольджи контролируют силу мышечного сокращения — напряжения. Идущие от этих рецепторов афферентные волокна относятся к группе Ib. На спиналь-ном уровне они через интернейроны вызывают торможение мотонейронов собственной мышцы и возбуждение мотонейронов мышцы-антагониста. Информация от мышечных рецепторов по восходящим путям спинного мозга поступает в высшие отделы ЦНС, включая кору большого мозга, и участвует в кинестезии. Таким образом, состояние мышцы контролируется двумя типами рецепторов – мышечными веретенами, передающими информацию о длине мышцы и скорости изменения этой длины, и сухожильным рецептором, который сигнализирует о силе мышечного сокращения. В отличие от мышечных веретён рецепторы Гольджи не имеют собственной эфферентной иннервации. Их импульсы направляются в спинной мозг к вставочным тормозным нейронам. Далее происходит переключение импульсов на а-мотонейроны, которые тормозятся, в результате чего мышца расслабляется. Это называется «аутогенным торможением». Все сухожильные рефлексы начинаются с рецепторов Гольджи. Коленный рефлекс — это искусственный (сухожильный) рефлекс. Он наступает при растяжении мышечного веретена в ответ на удар молоточком. Коленный рефлекс в отличие от других рефлексов не может наступать с рецепторов Гольджи, так как для их раздражения нужна большая сила, а у мышечного веретена порог возбуждения низкий. От рецепторов возбуждение поступает в Ib, а оттуда на четырёхглавую мышцу бедра, и нога в коленном суставе выпрямляется. Тельца Фатера — Почини представляют собой инкапсулированные нервные окончания, локализуются в глубоких слоях кожи, в сухожилиях и связках, реагируют на изменения давления, которое возникает при сокращении мышц и натяжении сухожилий и связок.
Рис. 1. Рефлексы растяжения моносинаптический (I, от мышечных веретён, приводит к сокращению той же мышцы) и полисинаптический (II) Моносинаптическая дуга. Ia–проприоцептивные нервные волокна, отходящие от мышечного веретена, входят в задний корешок спинного мозга и сразу направляются в передний рог, где и образуют синапсы с a‑мотонейронами, посылающими сигналы к мышце. Полисинаптическая дуга дополнительно включает вставочный нейрон. На рис.1. - (II) представлена дуга тормозного рефлекса, возникающего при растяжении сухожильных рецепторов Гольджи. Проводниковый отдел. Чувствительность кожи и ощущение движения обусловлены проведением в мозг сигналов от рецепторов по двум основным путям (трактам): леминисковому и спинно-таламическому, значительно различающимся по своим морфологическим и функциональным свойствам. Существует и третий путь — латеральный тракт Морина, близкий по ряду характеристик к леминисковой системе. Леминисковый путь на всех уровнях состоит из относительно толстых и быстропроводящих миелинизированных нервных волоконПервые нейроны этого пути находятся в спинномозговом узле, их аксоны в составе задних столбов восходят к тонкому (ядро Голля) и клиновидному (ядро Бурдаха) ядрам продолговатого мозга, где сигналы передаются на вторые нейроны леминискового пути. Часть волокон, в основном несущих сигналы от суставных рецепторов, оканчивается на мотонейронах сегментарного спинального уровня. Проприорецептивная чувствительность передается в спинном мозге также по дорсальному спинно-мозжечковому, спинно-цервикальному и некоторым другим путям. В продолговатом мозге в клиновидном ядре — вторые нейроны проприорецептивной чувствительности. Аксоны этих нейронов образуют медиальную петлю и после перекреста на уровне олив направляются в специфические ядра таламуса — вентробазальный ядерный комплекс. В этих ядрах концентрируются третьи нейроны леминискового пути. Их аксоны направляются в соматосенсорную зону коры большого мозга. Спинно-таламический путь. Этот путь значительно отличается от лемнискового. Его первые нейроны также расположены в спинномозговом узле, откуда они посылают в спинной мозг медленнопроводящие немиелинизированные нервные волокна. Эти нейроны имеют большие рецептивные поля, иногда включающие значительную часть кожной поверхности. Вторые нейроны данного пути локализуются в сером веществе спинного мозга, а их аксоны в составе восходящего спинно-таламического пути направляются после перекреста на спинальном уровне в вентробазальный ядерный комплекс таламуса (дифференцированные проекции), а также в вентральные неспецифические ядра таламуса, внутреннее коленчатое тело, ядра ствола мозга и гипоталамус. Локализованные в этих ядрах третьи нейроны спинно-таламического пути лишь частично дают проекции в соматосенсорную зону коры.
|
||||
|
|
Последнее изменение этой страницы: 2016-08-12; просмотров: 251; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.137.177.116 (0.007 с.) |