
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Мембранный потенциал покоя в нервной клетке.Содержание книги
Поиск на нашем сайте
В нейроне и в других клетках 1. мембрана в 75-100 раз более проницаема для К+ и С1-, чем для Na+, и практически не проницаема для внутриклеточных анионов, 2. концентрация К+ и непроникающих органических анионов намного больше внутри, чем снаружи, 3. концентрация Nа+ и С1- намного больше снаружи, чем внутри, 4. К+/Na+-АТФаза постоянно поддерживает трансмебранное распределение ионов К+ и Na+. Концентрации ионов в клетке и снаружи для нейрона млекопитающих показаны в таблице 1. Таблица 1
У каждого вида ионов концентрации по обе стороны мембраны неодинаковы. Химический градиент стремится переместить ионы через мембрану в сторону меньшей концентрации. Однако для катионов Nа+ и Са++ мембрана почти не проницаема и их свободный вход невозможен, а внутриклеточные анионы в нейроне велики и также не проходят через мембрану. Это в основном отрицательно заряженные белки и аминокислоты. Для калия проницаемость мембраны максимальна. Поскольку ионы К+ присутствуют в клетке в большем количестве, чем в межклеточной жидкости, они стремятся диффундировать из клетки в окружающую среду по градиенту концентрации. К+ выходит без сопровождения органических анионов, движущихся за ним в цитоплазме, но остающихся в клетке под мембраной и удерживающих катионы на внешней поверхности мембраны. В результате снаружи будет увеличиваться положительный заряд, а внутри – отрицательный. Так как противоположные заряды притягивают друг друга, положительные заряды и отрицательные заряды будут располагаться с каждой стороны у поверхности мембраны (рис.2). Разделение зарядов ведет к появлению трансмембранной разности потенциалов. Формирующаяся электрическая сила противодействует силе концентрационного градиента и препятствует дальнейшему выходу ионов К+. При равенстве электрических сил, определяемых зарядами на мембране, и противоположно направленных химических сил, связанных с градиентом концентрации, выход ионов К+ прекратится (вероятностное движение ионов К+ в клетку и из клетки уравновесится и общий трансмембранный ток К+ станет равным нулю). Величина мембранного потенциала, которая предотвращает движение ионов через мембрану в любом направлении, называется равновесным потенциалом (потенциалом Нернста или диффузионным потенциалом). Это энергия электрического потенциала, равная энергии концентрационного градиента (т.е. химического потенциала). Величина этого потенциала зависит от разницы концентраций – чем больше разница концентраций, тем больше стремление иона диффундировать в одном направлении и тем больше должен быть равновесный потенциал. Равновесный потенциал рассчитывается по следующей формуле Нернста
где Е – равновесный электрический потенциал; R – универсальная газовая постоянная; T – температура по шкале Кельвина; z – валентность иона; F – число Фарадея C – концентрация иона, в данном случае К + Уравнение Нернста может быть упрощено заменой натурального логарифма десятичным ln y =2,303 lg y и подстановкой значений констант F, R и T =370C (3100К). При использовании этой формулы предполагается, что потенциал снаружи равен нулю, а потенциал Нернста рассматривается для внутренней стороны мембраны и берется со знаком минус
Таким образом, чтобы создать электрический отрицательный потенциал внутри клетки достаточно только переместить катионы наружу и возникнет электрический диполь. Следует заметить, что достаточно предельно малого количества ионов, вынесенных из клетки, чтобы создать мембранный потенциал в –90 мВ внутри нервного волокна: примерно 1/5.000.000-1/100.000.000 от общего количество катионов должны быть перемещены из клетки наружу. Соответственно, такое же малое количество положительных ионов, движущихся из среды в клетку, могут изменить потенциал от –90 мВ до +35 мВ за 1/10000с. (см. далее Потенциал действия). В целом (и это важнейший фактор!) электрические процессы на мембране не приводят к измеримым химическими методами изменениям в концентрации ионов и потере макроскопической электронейтральности. В таблице 1 даны значения потенциала Нернста для некоторых ионов в нейроне млекопитающих. Так как в нейроне отношение концентраций К+ снаружи и внутри примерно 1 к 35, то расчетный потенциал Нернста для крупного нервного волокна, который находится в состоянии покоя, равен примерно –90 мВ. Это означает, что потенциал внутри клетки примерно на -90 мВ более негативный, чем потенциал в экстраклеточном растворе по отношению к цитоплазме. Следовательно, если мембранный потенциал покоя вызывается только током К+ через мембрану, то он должен быть примерно –90 мВ (см. далее). Уравнение Нернста может быть использовано для указания направления движения ионов через мембрану (рассмотрим пример для движения иона К+): 1. Если величина мембранного потенциала равна величине равновесного потенциала для какого-либо иона, то движение этого иона через мембрану (электрический ток) равно нулю.(в клетке при МП=-90 мВ ионы К+ не перемещаются через мембрану) 2. Если мембранный потенциал одинаков по заряду (положительный или отрицательный), но больше по величине, чем равновесный потенциал для какого-либо иона, то электрическая сила больше силы концентрационного градиента и ион будет двигаться по направлению электрической силы против градиента концентрации (если МП клетки увеличить до -100 мВ и более, ионы К+ начнут двигаться в клетку против градиента концентрации). 3. Если мембранный потенциал одинаков по заряду (положительный или отрицательный), но меньше по величине, чем равновесный потенциал для какого-либо иона, то сила концентрационного градиента больше, чем электрическая сила и ион будет двигаться по концентрационному градиенту (если МП в клетке меньше чем -90 мВ, то ионы К+ выходят из клетки по химическому градиенту). 4. Если мембранный потенциал противоположен по заряду равновесному потенциалу, то электрическая сила и сила концентрационного градиента для иона направлены в одну сторону. В этих условиях ион никогда не может быть в равновесии и движется в направлении, определяемом обеими силами одновременно (если МП клетки меняет знак на положительный, то ионы К+ выходят из клетки) Мембрана абсолютно проницаема для ионов С1- и его равновесный потенциал, рассчитанный по уравнению Нернста, примерно равен равновесному потенциалу для ионов К+, Между концентрациями проникающих ионов возникает следующее соотношение Часто возникает вопрос: «А не является ли ион С1 доминирующим в образовании мембранного потенциала?» Ион С1- не является основным в образовании МП, так как К+/Na+-АТФаза (см. далее) создает и поддерживает градиент для К+ и Na+, но не для С1-. В большинстве мембран нет активного транспорта ионов С1- . В таких клетках мембранный потенциал, создаваемый выходящим ионом К+ и остающимися в клетка анионами, допускает вход ионов С1- через мембрану по градиенту концентрации и в направлении отрицательно заряженной стороны только до тех пор, пока сила концентрационного градиента (количество ионов С1- снаружи больше, чем внутри) больше силы потенциала, удерживающего ионы С1- за пределами клетки. Следовательно, ионы С1- равновесно распределены в соответствии с мембранным потенциалом и практически не изменяют его величину в большинстве клеток. Если же распределение ионов С1- не соответствует мембранному потенциалу клетки, значит работает «активный» мембранный транспорт этого иона (например, Cl-/HCO3- обмен). В нейронах ионы С1- играют минимальную роль в установлении мембранного потенциала, но в некоторых случаях повышение проницаемости мембраны для иона С1, определяемое состоянием ионных каналов, это эффективный путь стабилизации МП, измененного каким-либо транспортом К+ или Na+. Изменение проницаемости для С1- является одним из способов воздействия на синаптическую передачу. Снижение проницаемости мембраны для С1- облегчает смещение МП от равновесного для клетки. Некоторые патологические миотонии, при которых наблюдается гипервозбудимость скелетных мышц, вызваны снижением проницаемости мембраны для ионов С1- вследствие врожденного нарушения хлорных каналов и, соответственно, снижением стабильности мембранного потенциала мышечных волокон. Согласно уравнению Нернста, равновесие устанавливается только для проникающих через мембрану ионов, но не для непроникающего аниона. В этом случае суммарная концентрация ионов в клетке всегда будет больше, чем в среде. Возникает осмотический дисбаланс, который приводит к перемещению внутрь клетки воды, стремящейся выровнять осмотические давления по обе стороны мембраны. Это важнейшее следствие равновесия Доннана! Если мембрана клетки легко растяжима и объем ее может меняться, возникают проблемы самого существования этой клетки: увеличивается объем, уменьшается концентрация непроникающих ионов внутри, уменьшается мембранный потенциал, выравниваются концентрации проникающих ионов по обе стороны мембраны и клетка разрушается. Стабильное существование клетки, в которой неравно распределены проникающие через мембрану ионы и существует разность потенциалов, возможно только при наличии в среде не проникающего в клетку компонента (заряженного или не заряженного) в концентрации, выравнивающей осмотические давления в клетке и в среде. В организме осмотический дисбаланс между цитоплазмой и внеклеточной средой не возникает исключительно благодаря наличию осмотически активного иона Na+ за пределами клетки. Наличие этого иона не только поддерживает осмотическое равновесие, но и уменьшает величину мембранного потенциала, поскольку суммарный потенциал мембраны любой клетки складывается из распределения всех проникающих ионов и определяется уравнением Гольдмана
Если мембрана высоко проницаема для К+, но слабо проницаема для Na+, то диффузия К+ вносит больший вклад в величину мембранного потенциала, чем диффузия Na+. В нормальном нервном волокне проницаемость мембраны для К+ примерно в 100 раз больше, чем для натрия. Используя эти значения в уравнении Гольдмана, получим потенциал примерно /–70/-/-85/ мВ, который меньше калиевого равновесного потенциала в -90 мВ, хотя и близок к нему. Уравнение Гольдмана (или Гольдмана-Ходжкина-Катца) показывает, что мембранный потенциал является средним взвешенным равновесным потенциалом всех ионов, которые могут проходить через мембрану и не является равновесным ни для одного из них. Разница между величиной мембранного потенциала и равновесным потенциалом для каждого конкретного иона является движущей силой для перемещения этого иона через мембрану. По закону Ома движение каждого иона через мембрану зависит от проводимости для иона (g) и электродвижущей силы (напряжения, разности потенциалов). Для Na+, K+, Cl-
INa –величина тока для соответствующего иона, Еm –мембранный потенциал, ЕNa- - равновесный потенциал для данного иона. Таким образом, постоянный мембранный потенциал (потенциал покоя) является примером динамического постоянства и поддерживается равенством Величина мембранного потенциала покоя способствует утечке К+ из клетки и - одновременно движению Nа+ в клетку однонаправленно действующими электрической силой и силой концентрационного градиента. Входящий Na+ еще больше повышает осмотическое давление внутри клетки и ее объем за счет поступающей воды, уменьшает величину мембранного потенциала и в итоге клетка может не просто потерять потенциал, но и полностью разрушиться из-за осмотического шока. В таких условиях нормальная клетка может существовать только поддерживая ионные концентрации и защищаясь от поступления воды дополнительным – насосным - током, создаваемым работой Na+/К+-АТФазы, которая, используя независимый источник энергии (энергию АТФ), сохраняет внутри клетки низкую концентрацию ионов натрия, выводит его из состояния электрохимического равновесия и поддерживает осмотическое равенство между цитоплазмой и средой. К+/Na+-АТФаза непрерывно перемещает 3 катиона Na+ наружу и одновременно 2 катиона К+ внутрь. Именно она создает и постоянно поддерживает высокую внутриклеточную концентрацию К+ и высокую внеклеточную концентрацию ионов Na+. Выход Na+ уменьшает осмотическое давление внутри клетки и увеличивает осмотическое давление внеклеточной жидкости. Для перемещения ионов Na+ против электрохимического градиента затрачивается энергия расщепления молекулы АТФ. Неэквивалентным обменом ионов К+/Na+-АТФаза создает дополнительный отрицательный потенциал в клетке от -5 до -20 мВ в зависимости от сопротивления мембраны. Основным фактором, активирующим работу К+/Nа+-АТФазы, является повышение концентрации внутриклеточного натрия. Она не чувствительна ни к концентрации внеклеточного натрия, ни к величине МП.
Вопросы для самоконтроля 1. Что такое мембранный потенциал? 2. Как измерить величину мембранного потенциала? 3. Каковы ионные механизмы потенциала покоя? 4. Что такое диффузионный потенциал? 5. Расскажите об электродвижущей силе для ионов и ионных токах. 6. Что такое равновесный потенциал? Напишите уравнение Нернста. 7. Что такое осмотическое неравенство между клеткой и средой? Чем оно опасно?Как поддерживается осмотический баланс? 8. Какие ионы определяют мембранный потенциал покоя нейрона? Напишите уравнение Гольдмана и объясните роль каждого иона. 9. Почему существуют постоянные токи утечки ионов К+ и Na+ через мембрану? Как они могут изменить мембранный потенциал? 10. Какую роль играет Na+/K+-АТФаза в поддержании мембранного потенциала? 11. Величина ПП даже при отсутствии воздействия на клетку или волокно испытывает некоторые колебания. С чем это связано? 12. В ходе измерения величины ПП микроэлектродным методом (с использованием стеклянногомикроэлектрода-пипетки, заполненного 3М раствором КС1) она со временем начинает уменьшаться. В чем причина этого явления?
|
||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-14; просмотров: 359; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.17.174.204 (0.007 с.) |

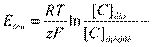
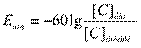 ;
;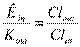 , называемое уравнением Гиббса-Доннана. В клетке значительно преобладают ионы К, удерживаемые отрицательным зарядом органических анионов, снаружи - ионы Cl- , которые этим же отрицательным зарядом анионов удерживаются за пределами клетки. При этом оба иона находятся в состоянии электрохимического равновесия.
, называемое уравнением Гиббса-Доннана. В клетке значительно преобладают ионы К, удерживаемые отрицательным зарядом органических анионов, снаружи - ионы Cl- , которые этим же отрицательным зарядом анионов удерживаются за пределами клетки. При этом оба иона находятся в состоянии электрохимического равновесия.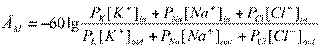
 ,
, 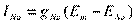 ,
,  , где
, где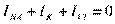 , в котором суммарный трансмембранный ток равен нулю.
, в котором суммарный трансмембранный ток равен нулю.


