Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
НЭ - неэнзиматические факторы.Содержание книги
Поиск на нашем сайте
_____________________________ ше, комбинацию микрополостного и мембранного пищеварения. Следует заметить, что у некоторых насекомых первая стадия пищеварения протекает не только вне пищеварительной полости, но даже за пределами организма. Однако это только кажущееся противоречие. Хотя такие насекомые начинают переваривать свою жертву еще до поглощения, вводя свои ферменты в ее тело, ясно, что это - определенная модификация внеклеточного полостного пищеварения. Существуют и еще кажущиеся исключения. Так, многие кишечные па-
2* разиты потеряли способность секретировать пищеварительные ферменты. Однако внеклеточное пищеварение у них происходит за счет ферментов хозяина. Более того, у ряда паразитов часто адсорбируются ферменты хозяина для реализации промежуточных этапов мембранного пищеварения (обзоры: Уголев, 1985; Уголев и др., 1985). В большинстве случаев усвоение пищевых веществ требует участия симбионтов - бактерий и простейших (см. 1.3.5). Таким путем ряду организмов удалось приспособиться к потреблению целлюлозы, шерсти, хитина и других материалов, не гидролизуемых собственными ферментами. Однако и здесь взаимодействуют известные типы пищеварения у макроорганизма и его симбионтов.
Симбионтное пищеварение и питание
Особое место среди других механизмов переваривания пищи занимает симбионтное пищеварение, т. е. пищеварение за счет микроорганизмов желудочно-кишечного тракта. Оно присуще почти всем многоклеточным организмам и отчетливо проявляется у растительноядных жвачных. Симбионтное пищеварение характерно и для других животных, подобных истинным жвачным, например для верблюда, ламы и др. Этот тип пищеварения широко распространен также у беспозвоночных, например у насекомых. Симбионтное пищеварение подробно охарактеризовано в ряде капитальных обзоров (Prosser, Brown, 1967; Сравнительная физиология..., 1977; Schmidt-Nielsen, 1982; Martin, 1989; Williams, 1989,, и др.). Как правило, пищеварительный тракт макроорганизма заселен бактериями и простейшими, которые частично или полностью снабжают организм хозяина необходимыми органическими веществами, в том числе витаминами, незаменимыми аминокислотами и др. Симбионтами некоторых животных. например моллюсков, актиний, кораллов, могут быть также водоросли - зооксантеллы и зоохлореллы, поставляющие пищевые вещества хозяину. Интересно, что у хищных насекомых симбионты отсутствуют, тогда как у тех, кто потребляет сок растений, симбионтное пищеварение играет весьма важную роль.
Анализ различных форм и модификаций симбионтного пищеварения позволил выделить в нем два фундаментальных биологических механизма: 1) первичные пищевые вещества разрушаются ферментами бактерий и
простейших, а образующиеся продукты гидролиза ис-пользуются преимущественно организмом хозяина; 2) бактнрии и простейшие не только разрушают первич-иные пищевые продукты, но и утилизируют их, тогда как хозяин поглощает вторичную пищу, состоящую из структур симбионтов. Первый механизм точнее было бы назвать симбионтным пищеварением, а второй — симбионтным питанием в сочетании с симбионтным пищеварением. Животные со специализированными симбионтами, способными использовать метан и серу, оказались в начале трофической цепи в биологических изолятах на дне океанов, где наблюдалась высокая вулканическая активность. В этом случае существуют выходы метана или сероводорода, которые служат внешними источниками энергии подобно свету. Симбионтные взаимодействия широки и разнообразны. Например, приспособление жвачных к использованию грубых растительных кормов во всех случаях связано с действием на эти субстраты симбионтов. Можно не считать преувеличением, что существование таких организмов в основном базируется на использовании симбионтных процессов. При рассмотрении симбионтных питания и пищеварения взаимодействия бактериальной флоры и организма-хозяина анализируются обычно на примере человека и высших животных, преимущественно жвачных. Однако для понимания механизмов питания и пищеварения в естественных условиях, а также для формирования новых оптимальных искусственных условий целесообразно анализировать весь круг явлений, известных среди организмов. Нутритивные симбиозы известны уже у простейших, в которые вселяются бактерии и некоторые водоросли, обеспечивающие их продуктами фотосинтеза. Среди многоклеточных организмов уже у кишечнополостных обнаруживаются зоохлореллы и зооксантеллы, синтезирующие углеводы и снабжающие организм хозяина дополнительным кислородом.
Организмы, содержащие симбионты, более устойчивы к пищевому голоданию и недостатку кислорода. Можно думать, что во многих случаях быстрота адаптации и, следовательно, возможность изменения трофической ниши связаны не только, а иногда и не столько с изменением пищеварительных характеристик самого хозяина, сколько с изменением ферментных характеристик микрофлоры, населяющей желудочно-кишечный тракт (см. гл. 3). Много ярких примеров этому мы видим у насекомых. Так, питание шерстью или шелком зависит от формирования специфической кишечной флоры и по своему механизму является адаптацией на уровне бактериальной флоры. Следовательно, необходима разгадка того, каким образом организм хозяина культивирует нужный тип микрофлоры и затем передает потомству иногда чистую или почти чистую и хорошо стабилизированную культуру бактерий. Более того, образование экзотических трофических ниш и заполнение трофических пустот также в ряде случаев связаны с приобретением и культивированием соответствующих симбионтов. Нельзя исключить, что в составе нормальной флоры желудочно-кишечного тракта всегда присутствуют «случайные» загрязняющие бактерии, способные утилизировать и расщеплять нехарактерные для данной популяции пищевые субстраты. Таким путем, вероятно, происходят эволюция питания и приспособление организмов к новым трофическим нишам. Кроме того, у многоклеточных организмов в пределах пищеварительного аппарата могут существовать своеобразные трофические цепи, что обеспечивает многие важнейшие эволюционные и экологические перестройки. Например, термиты, питающиеся преимущественно древесиной, обеспечивают свои потребности в белке за счет включения в кишечную флору азотфиксирующих бактерий. По-видимому, благодаря присутствию в желудочно-кишечном тракте азотфиксирующих бактерий действительно возможны подлинная фиксация атмосферного азота и включение его в биологический цикл, минуя промежуточные звенья растительной трофической цепи. Этот пример является многообещающим и демонстрирует недостаточно изученную, но исключительно важную сторону эволюции процессов питания и пищеварения в животном мире.
Индуцированный аутолиз
Ранее нами было высказано предположение о важной роли так называемого индуцированного аутолиза в ассимиляции пищи. Согласно нашей гипотезе, этот тип пищеварения, при котором объект питания в естественных условиях обеспечивает собственное переваривание и усвоение, является наиболее древним механизмом начальной переработки пищевых веществ у многоклеточных организмов. Индуцированный аутолиз реализуется собственными ферментами пищевого объекта, а пищеварительные ферменты хозяина создают благоприятные условия для их действия. Однако до последнего времени считалось, что аутолиз играет незначительную роль в общем пищеварительном процессе, так как начальные этапы пищеварения осуществляются в основном в полости желудочно-кишечного тракта высших организмов секретируемыми Ферментами хозяина, а промежуточные и заключительные - в результате мембранного пищеварения. Таким образом, было важно установить относительную роль ферментов хозяина и пищевого объекта в начальных этапах гидролиза пищевых веществ при питании в естественных условиях. Особенно трудно было интерпретировать питание хищников, у которых происходит переваривание больших порций мяса, из-за чего отношение поверхность/масса мало и невыгодно для ферментов желудочного сока, действующих на поверхности пищевого объекта.
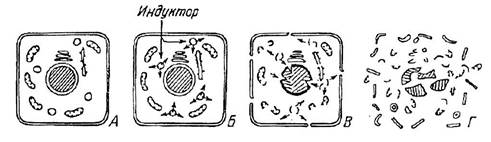
Индуцированный аутолиз был обнаружен нами около 20 лет назад, но описан лишь в 1984 г. (Уголев, Цветкова, 1984). (Детально этот механизм освещен также э обзорах: Уголев, 1985, 1987а.) При индуцированном аутолизе жертва, или, шире говоря, объект питания, обеспечивает свое собственное переваривание. Так, например, происходит, если удав заглатывает кролика. До последнего времени было неясно, каким образом проглоченная целиком жертва переваривается удавом. Действительно, поверхность контакта жертвы с ферментами желудочного сока хищника сравнительно мала, так как пищевой объект не раздроблен. Оказалось, однако, что раньше, чем ферменты хищника гидролизуют структуры жертвы, начиная с ее поверхности, организм этой жертвы будет аутолизироваться за счет индуцированного аутолиза. Индуцированный аутолиз был исследован нами в модельных экспериментах, названных «маленький искусственный удав>. В прозрачную камеру, заполненную желудочным соком человека, лошади или собаки, помещались «сырая» лягушка и лягушка после недолгой термической обработки. В первые несколько часов гидролиз сухожилий термически обработанной лягушки шел быстрее, чем «сырой», что служило подтверждением общепринятых взглядов. Однако в последующие 2-3 дня «сырая» лягушка полностью растворялась, тогда как структуры термически обработанной лягушки в значительной мере сохранялись. Таким образом, в этих экспериментах наряду с доказательством существования индуцированного аутолиза было продемонстрировано, что нативные белки гидролизуются быстрее денатурированных. В дальнейшем мы детально исследовали механизм переваривания нативных тканей желудочным соком. Выяснилось, что суть этого механизма состоит в следующем. Кислый желудочный сок хищника индуцирует самопереваривание жертвы ее же ферментами. Под действием этого сока происходят разрушение лизосом и выход в клетку многочисленных лизосомальных гидролаз, разрушающих все структуры клетки при рН 3.5-5.5. Итак, организм-ассимилятор индуцирует расщепление структур пищевого объекта ферментами последнего, активируя их и создавая оптимальные условия среды, в том числе рН. В соответствии с классическими представлениями о механизмах переваривания пищевого объекта, ферменты пищеварительных соков осуществляют свой эффект только поверхностно. Скорость диффузии гидролаз внутрь пищевого объекта даже при низком диффузионном сопротивлении мембраны лимитирована их сравнительно большой молекулярной массой, тогда как скорость диффузии протона примерно на 3-4 порядка больше. При выходе гидролаз из лизосом под влиянием сдвига рН возникает множество центров гидролиза в каждой клетке (рис. 1.4), что создает практически тотальное расщепление ткани. Следует добавить, что в кислых секретах организма-ассимилятора содержатся главным образом протеазы, тогда как ферментный спектр лизосом практически универсален. Однако в ассимилируемых объектах имеются также структуры (например, белки соединительной ткани, жировые депо, в тканях растений - полисахаридные депо), лишенные лизосом и
_____________________________
Рис. 1.4. Упрощенная концептуальная схема внутриклеточных процессов при индуцированном аутолизе (по: Уголев, 1985).
|
||||||||
|
|
Последнее изменение этой страницы: 2016-06-26; просмотров: 387; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.141.19.115 (0.012 с.) |