Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Реакция диких копытных на хищных: – инстинкт или традиция.Содержание книги
Поиск на нашем сайте
Знание поведенческих взаимоотношений хищника – жертвы приобретает особое значение тогда, когда происходит восстановление находящихся на грани исчезновения или исчезнувших в природе видов, независимо от того, хищник это или копытное. Когда речь идет о защитной реакции копытного по отношению к хищнику, подразумевается тот поведенческий комплекс, который выражается в убегании, вокализации и в поведении, непосредственно связанном с охраной себя или потомства. Вальтер (Walter, 1970) показал, что «дистанция бегства» газели Томпсона – Gasella tomsoni (подразумевается то минимальное расстояние между хищником и потенциальной жертвой, при сокращении которого она убегает от хищника) зависит от пола, возраста и ранга индивида. Интересно, что у взрослых самцов эта дистанция намного меньше, чем у самок. У самок же – приблизительно такая же, как у молодых самцов. У самцов, имеющих пары, дистанция бегства меньше, чем у самцов, не имеющих таковых, а у последних, в свою очередь, меньше, чем у не территориальных самцов, у которых дистанция убегания больше, чем у самок. Автор считает, что внутрипопуляционная дифференциация дистанции бегства является, в первую очередь, средством поддержания социальной структуры популяции и только потом – адаптацией, позволяющей избежать нападения хищника. Представленная здесь интерпретация автора интересна сама по себе и полностью приемлема. Но мы хотим обсудить те стороны проблемы, которых автор не коснулся. Исходя из цитированного материала, можно сделать еще один вывод: меньшая «дистанция бегства» доминантных индивидов обуславливает еще и большую вероятность гибели их (производителей) от хищника. Это, в свою очередь, обеспечивает смену доминантов и способствует сохранению их оптимального возраста (производителей), как это уже было показано раньше (Бадридзе и соавт., 1992). Дистанция бегства, как одна из пороговых форм защитного поведения, в свое время была описана Баскиным (1976). В его работе описана дистанция бегства некоторых видов копытных от человека. Как выясняется, она различна у разных видов. Из приведенной работы можно сделать вывод – дистанция бегства зависит от потенциальной скорости передвижения (или бега) вида и характерного для него психологического типа. Особого внимания заслуживает работа Шаллера (Schaller, 1972), в которой автор показал, что дистанция бегства зебры от хищника зависит от вида самого хищника. Так, например, эта дистанция при убегании от пятнистой гиены равна 9 м., от гиеновой собаки – 18 м., льва – 45 м., а если в стаде находятся жеребята, то и 90 м. Следовательно, дистанция бегства от хищника прямо пропорциональна потенциальной скорости последнего; в то же время учитываются скоростные возможности находящихся в стаде жеребят. Несмотря на то, что приведенными трудами ограничиваются данные по рассматриваемой теме, они полностью отражают сущность указанного поведенческого феномена. Однако они, к сожалению, не отвечают на поставленный нами вопрос: является ли защитная реакция копытных инстинктом на хищника или приобретенной формой поведения, ставшей традицией? Поэтому нашей целью было изучение защитного поведения (поведенческого иммунитета) копытных по отношению к хищнику, природы данного поведения и механизма его формирования. Изучение этого вопроса поможет определить оптимальный темп реинтродукции хищника.
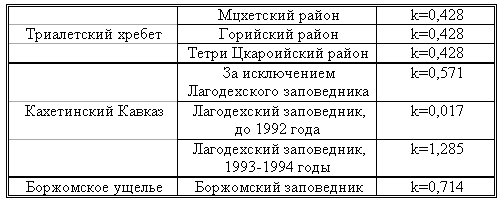
Материал и методика Наблюдения и эксперименты проводили в природных условиях на 860 кавказских благородных оленях (Cervus elaphus marsal) с оленятами и на 300 дагестанских турах (Capra cilindricornis) с козлятами. Изучали реакцию диких копытных на человека и на хищных. Для изучения защитной реакции на человека наблюдения и эксперименты проводили на территории Грузии, в регионах, подвергшихся наибольшему прессу браконьерства (Триалетский хребет – Мцхетский, Горийский, Тетрицкаройский районы, Боржомский заповедник, Кахетинская часть Большого Кавказа, включая Лагодехский заповедник). Кроме этого, специальные наблюдения проводили в Лагодехском заповеднике в период интенсивного браконьерства (1992-1994 годы). Для определения уровня браконьерства систематически считали услышанные выстрелы и определяли направление, откуда они доносились. одним фактом браконьерства считали случай, когда выстрелы раздавались из одного и того же места, независимо от их количества. Для каждых 7 дней вычисляли коэффициент уровня браконьерства k=n/7, где n-число ежедневных случаев браконьерства. Исходя из полученного коэффициента, устанавливали влияние уровня браконьерства на дистанцию бегства копытных. Изучение реакции диких копытных на человека велось двумя методами: I – приближались открыто; II – приближались скрытно на возможно минимальную дистанцию, после чего появлялись перед животными. В обоих случаях описывали поведенческую реакцию копытных и их детенышей, вокализацию и дистанцию бегства. При изучении реакции копытных на хищников наблюдения и эксперименты проводили по отдельности, в регионах, где на протяжении нескольких лет не было волков (территории Мцхетского и Горийского районов на отрезке Триалетского хребта), а также в регионах, где волки обитали постоянно (Боржомский и Лагодехский районы). В Триалети наблюдения вели за дистанцией бегства копытных от реинтродуцированных нами волков. В других же районах – подкрадываясь на 200-300 метров к группе копытных, выпускали выращенного в неволе волка с намордником (из 53 экспериментов в 20 случаях волк не умел охотиться) и наблюдали за их реакциями. Полученные результаты и их обсуждение Прежде чем изложить материалы, полученные в результате наблюдения за животными, целесообразно привести таблицу коэффициентов, отражающих уровень браконьерства в изученных нами регионах (таблица 16).
Таблица 16. Коэффициент уровня браконьерства в различных регионах Грузии.
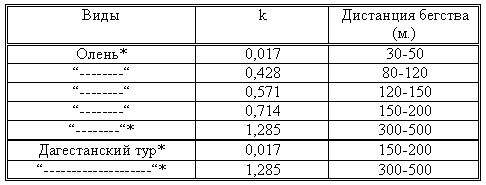
Изучение дистанции бегства от нас диких копытных показало, что ее показатель находится в прямой зависимости от коэффициента (к) уровня браконьерства. Так, например, дистанция бегства от нас была минимальной в тех районах, где к=0, 017 (кавказский олень – 30-50м), а там, где коэффициент браконьерства был максимальным (к=1, 285) соответственно увеличивалась и дистанция бегства (кавказский олень – 300-500). Детально эти данные приведены в таблице 17.
Таблица 17. Влияние уровня браконьерства на дистанцию бегства копытных от человека. * – одни и те же популяции.
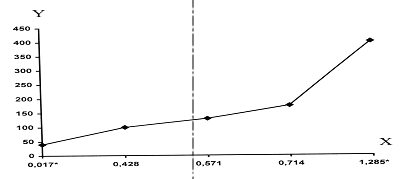
Данные об изменении уровня браконьерства в одном и том же регионе получены нами только в Лагодехском заповеднике. Однако и их достаточно, чтобы было очевидным влияние этого фактора на дистанцию бегства представителей одной и той же популяции (рис. 17). Полученный материал однозначно указывает на то, что дистанция бегства не является врожденной. Она может изменяться пропорционально изменению пресса браконьерства на копытных. Исходя из этого, об уровне браконьерства можно судить по дистанции бегства копытных от человека. По нашему мнению, это имеет большое практическое значение для оперативного выявления ситуации в том или ином районе.
Рисунок 17. Зависимость дистанции бегства Кавказского благородного оленя от уровня браконьерства. X – коэффициент браконьерства; Y – средняя дистанция бегства. * – одни и те же популяции.
Следует отметить тот факт, что дистанция бегства существующих в группе молодых индивидов и взрослых животных одинакова. Мы подчеркиваем, существующих в группе, т.к. дистанция бегства молодых может измениться после того, как они выйдут из-под опеки родителей и начнут самостоятельную жизнь. Полученный нами материал говорит и о том, что дистанция бегства взрослых индивидов меняется в зависимости от индивидуального (или группового) опыта. Потомству же опыт передается путем опосредованного обучения. Исходя из этого, следует подчеркнуть, что дистанция бегства индивида не является врожденной и передается следующему поколению за счет врожденной способности к подражанию. Последнее же – один из значительных механизмов обучения. Эксперименты, проведенные при подкрадывании к диким копытным на возможно минимальную дистанцию, показали, что самки оленей и туров с детенышами при внезапном появлении человека на близком расстоянии издают специфические звуки (олени – резкий лающий звук, туры – свист). Вслед за этим убегают взрослые особи; детеныши же, как правило, убегают сразу при звуке тревоги. В некоторых случаях (n=27) детеныши убегали только вместе с родителями. При повторном подкрадывании к этим индивидам детеныши убегали не вместе с родителями, а на фоне звуков тревоги, издаваемых родителями. Описанные факты указывают на то, что реакция убегания детенышей формируется за счет одноразового обучения. Для этой формы обучения решающее значение имеет врожденная реакция на издаваемый родителем звук тревоги и сопряженная с этим звуком поведенческая реакция того же родителя. Следует подчеркнуть и то, что этот звук является для детеныша безусловным раздражителем. Именно сопряжение этого раздражителя с появлением человека обуславливает формирование защитного поведения детенышей за счет одноразового обучения. При сопоставлении материала, полученного нами на волках и копытных, стало очевидным, что и у хищных, и у копытных в процессе постнатального онтогенеза описанная поведенческая реакция должна формироваться по одному и тому же принципу. Можно предположить, что поведенческая реакция родителей формирует традицию конкретной формы защитного поведения потомства и ее передачу от поколения к поколению путем опосредованного обучения. На основе вышесказанного можно сделать вывод, что ситуационное выражение и интенсивность защитной формы в зависимости от поведения не является врожденной и поэтому может меняться в зависимости от состояния окружающей среды. Этим обеспечивается формирование оптимальной формы поведения и ее интенсивности, и тем самым адаптация к условиям окружающей среды. Наблюдения за охотой подготовленных нами волков показали, что с момента реинтродукции в течение 6 месяцев дистанция бегства от них диких копытных возросла приблизительно в 6 раз. Изменение дистанции бегства показано на графике 18.
Рисунок 18. Изменение дистанции бегства косуль от реинтродуцированных волков. X – месяцы со дня реинтродукции; Y – дистанция бегства в метрах. Рисунок 19. Изменение процента успешной охоты реинтродуцированных волков. X – месяцы со дня реинтродукции; Y – процент успешной охоты.
Как видно из приведенного материала, между дистанцией бегства от хищника и успешностью охоты последнего существует отрицательная корреляция (рис. 19). Для формирования такой корреляции, по-видимому, большое значение имеет опыт, приобретенный «спасшейся» жертвой. Этот опыт использует не только она сама, но и передает его другим (включая потомство) путем опосредованного обучения. Интересно и то, что дистанция бегства копытных увеличивается и в том случае, когда индивиды наблюдают смерть своих собратьев. Поэтому обучение возможно не только за счет собственного опыта или при наблюдении за завершенной поведенческой реакцией других особей, но и тогда, когда другой индивид успевает осуществить лишь небольшой фрагмент защитного поведения и оно кончается неудачей. Исходя из этого, одноразовое сочетание внешних признаков врага (хищник, человек) и защитного поведения или его фрагмента у собрата достаточно для формирования адекватной защитной реакции животного – «наблюдателя». Сразу же после восстановления у копытных защитной реакции по отношению к волку в вышеуказанных регионах провели эксперименты с целью изучения их реакции на хищников, не имеющих опыта охоты. Эксперименты показали, что во всех 20 случаях у копытных развивалась такая же четко выраженная защитная реакция, как и на особей, имеющих опыт охоты. Это еще раз указывает на решающее значение видовых признаков хищника для формирования у копытных оборонительной реакции. Эксперименты, проведенные в регионах постоянного обитания волка, показали, что реакция диких копытных на волков четко дифференцирована. В частности, на имеющих опыт охоты хищников у копытных имеется ярко выраженная защитная реакция, тогда как они индифферентны по отношению к тем особям, которые не умеют охотится, т.е. не воспринимают копытных как потенциальную жертву. Такие индивиды могли находиться в непосредственной близости к оленям. Приведенный материал перекликается с работой Мичурина (1970), где он отмечает, что часто видел волков, которые разгуливали среди группы оленей. Расстояние между ними менялось от 30 до 100 метров. В другом случае волк сидел среди большой группы оленей на холме. Приведенный нами материал указывает на то, что при взаимодействии с хищником копытные приобретают опыт, который позволяет им различать не только его видоспецифичные черты, но и специфику поведения этого вида и, исходя из этого, и его «цель». Из всего вышесказанного можно сделать следующий вывод: реакция диких копытных на человека и хищников не является врожденной. Форма этой реакции и ее интенсивность могут изменяться соответственно уровню пресса на копытных. Приобретение опыта (обучение), может происходить с одного раза, не только во время непосредственно полученного «урока», но и методом наблюдения за реакцией других индивидов (опосредованное обучение). Для опосредованного обучения достаточно восприятия фрагмента реакции другого индивида, что полностью исключает необходимость приобретения собственного опыта, тем самым, значительно ускоряет процесс обучения и делает его значительно безопасным и, следовательно, имеет большое значение для выживания индивида. В случае стабильной ситуации в ареале, опыт, приобретенный индивидом, передается из поколения в поколение путем опосредованного обучения и приобретает форму поведенческой традиции. Как было показано, дистанция бегства копытных меняется в зависимости от уровня пресса браконьерства. Следовательно, сама дистанция бегства является безошибочным показателем уровня браконьерства. Наличие подобной зависимости позволяет оперативно определить ситуацию, существующую в том или ином регионе, что имеет важной практическое значение для контроля за уровнем браконьерства. Если интенсивность описанной защитной реакции не является врожденной, то она может угаснуть или ослабнуть в том случае, если в ареале копытного, по разным причинам, больше нет хищника. Наглядным примером этого служит дистанция бегства косули от реинтродуцированных нами волков. Как оказалось, эта поведенческая форма и ее интенсивность восстановимы, но в масштабе популяции для ее восстановления необходимо длительное время. Исходя из этого, в процессе восстановления вида хищника очень важно, какое количество животных реинтродуцируется одновременно. Реинтродукция одновременно большого числа хищных до восстановления защитного поведения копытных может отрицательно сказаться на их популяцию. Анализ приведенного материала позволяет сделать следующие выводы: 1. форма и интенсивность защитной реакции копытных по отношению к врагу (человеку или хищнику) подвержены сильной модификации в онтогенезе, вследствие чего возможны их изменения во времени в соответствии с существующей ситуацией; 2. в конкретной ситуации форма и интенсивность защитного поведения по отношению к врагу передается опосредованным путем из поколения в поколение; 3. у копытных, в случае отсутствия в их ареале хищников, через одно поколение гаснет оборонительная реакция по отношению к ним; 4. во избежание отрицательного воздействия на численность копытных, реинтродукция (судя по всему, и транслокация) восстанавливаемого вида хищников должна производиться небольшими группами и со значительными интервалами. Естественно, что интервал должен определяться, исходя из поведенческой и репродуктивной специфики копытных; 5. пресс браконьерства по отношению к копытным можно определять по дистанции их бегства от человека.
ЗАКЛЮЧЕНИЕ
Из полученного нами материала следует, что в процессе искусственного выкармливания новорожденных хищных млекопитающих, подготавливаемых к реинтродукции в природу, особое значение имеет период, когда животные питаются молоком. Необходимо отметить, что при сосании рожка животные должны иметь возможность полноценной реализации реакции «массажа лапками молочной железы». В противном случае, судя по всему, в определенных областях головного мозга формируются очаги с постоянно высоким уровнем активности, вследствие чего у животных развивается неконтролируемая гиперактивность передних конечностей и эмоциональная неуравновешенность. У подобных животных часто возникают фрустрации, и они постоянно провоцируют конфликты в группе. Во время искусственного выкармливания надо учесть необходимость обеспечения оптимального темпа поступления молока во время сосания. В том случае, если молоко поступает без приложения значительных сосательных усилий, у животных увеличивается естественный объем желудка, вследствие чего порог сенсорного насыщения повышается. В этих случаях животные насыщаются позже других, что приводит к возрастанию агрессивных взаимодействий на почве пищевой конкуренции. Вместе с тем, у этих животных не формируется поведение запасания пищи. Как было показано, поведение запасания пищи имеет большое значение для поддержания кормовой базы группы. Данный вид поведения приобретает особое значение во время выкармливания прибылого потомства, обеспечивая стабильность их кормовой базы. Присутствие в группе особей с не сформировавшимся поведением запасания не просто подрывает стабильность кормовой базы прибылых, но и может привести к значительному повышению агрессивных взаимодействий из-за пищи и, вследствие этого, к неритуализированности агрессивного поведения. Исходя из сказанного, животные с не сформировавшимся поведением запасания пищи не могут быть отобраны для реинтродукции в природу. Поведение ухода за детенышами формируется задолго до полового созревания, что дает великолепную возможность отбирать животных для подготовки к реинтродукции в природу с нужными качествами задолго до их половой зрелости. Чтобы сформировать характерный для вида максимальный уровень рассудочной деятельности, необходимо содержать животных в среде, предельно обогащенной крупными непрозрачными предметами. Животные, выросшие в подобных условиях, охотятся несравненно успешнее и легче адаптируются к новым условиям обитания после реинтродукции, чем выросшие в обедненной среде. Начиная с ювенильного периода, необходимо время от времени вывозить животных в естественную среду обитания, что в дальнейшем значительно ускоряет освоение ими выбранной для реинтродукции территории. Хищническое и охотничье поведение в процессе постнатального онтогенеза развивается поэтапно, причем каждый предыдущий этап является основой для последующего. Исходя из этого, подготовка крупных хищных к реинтродукции должна начинаться в раннем ювенильном возрасте. Во время этого процесса следует учитывать, что первая реакция на потенциальную жертву не связана с пищевой мотивацией. Эта связь образуется только после установления хищником факта съедобности жертвы, что происходит за счет непосредственного опыта или опосредованного обучения. Исходя из вышесказанного, в процессе подготовки к реинтродукции выращенных в неволе хищных для их нормального развития необходимо целенаправленное обучение животных тому, что потенциальная жертва является пищей. Только после того, как молодой хищник установит связь между потенциальной жертвой и пищей, можно переходить к его обучению охоте. Как было показано, этап формирования охотничьего поведения состоит из нескольких подэтапов. При этом полноценное развитие каждого подэтапа возможно только в так называемый «период чувствительности» для данного подэтапа. Из материала видно, что если хищнику в соответственный чувствительный период постнатального онтогенеза не дали возможности обучиться охоте, эта возможность полностью теряется или не развивается полноценно. В обоих случаях реинтродукция таких животных становится невозможной. В процессе подготовки к реинтродукции следует учесть и то, что хищникам, обладающим высокой социальностью, необходимо предоставить возможность групповой охоты. Это требуется для того, чтобы члены группы во время нападения на крупную жертву не только научились взаимосогласованным действиям, что значительно увеличивает успешность охоты, но и для развития навыков самой охоты. Все это формируется только путем накопления опыта методом проб и ошибок. К моменту реинтродукции животные должны быть хорошо знакомы с предназначенной для реинтродукции территорией, т.е. должны сформировать ее «ментальную карту». Это позволит им свободно ориентироваться на территории, что очень важно как для поиска объекта охоты, так и для выгона жертвы в желаемом для хищника направлении. К началу освоения территории, предназначенной для реинтродукции, необходимо начать выработку у хищных реакции избегания незнакомого человека и домашних животных. Это даст возможность исключить возникновение разного рода конфликтов между реинтродуцированным животным и местным населением. А если, к тому же, представители местного населения сами участвуют в этом процессе, то полностью меняется их отношение к реинтродуцированному хищнику. Это имеет одно из решающих значений для успешного восстановления вида. Как видно из полученного нами материала, реакция избегания домашних животных не вырабатывается генерализованно. Ее следует выработать у хищника в отношении каждого вида, независимо от породы. Этот принцип выработки реакции, по нашему мнению, выгоден и тем, что дает возможность выработки реакции избегания и в отношении тех видов диких животных, которые находятся на грани исчезновения. А это даст возможность восстановить вид хищника без его отрицательного воздействия на охраняемый вид. Было показано, что форма защитного поведения копытных по отношению к врагу (человеку, хищнику) и его интенсивность не являются врожденными (жестко запрограммированными), поэтому возможно их изменение во времени в соответствии с существующей ситуацией. Исходя из этого, в процессе восстановления вида хищника, во избежание отрицательного воздействия на численность жертвы необходимо реинтродуцировать их небольшими группами. Интервалы между реинтродукциями, по нашему мнению, должны соответствовать возрасту полового созревания данного вида, что будет достаточным для восстановления поведенческого иммунитета к хищнику у различных видов «жертвы». В заключение можно сказать, что, учитывая видовую специфику высшей нервной деятельности, разработанный нами метод можно применить при реинтродукции любого вида крупных хищных млекопитающих. А метод освоения территории и выработки реакции избегания незнакомых людей – и при реинтродукции копытных животных.
ОБЩИЕ ВЫВОДЫ
1. При искусственном выкармливании хищных млекопитающих особое значение для нормального формирования их поведения имеет неонатальный период, который в значительной мере определяет дальнейшее развитие поведения в пределах нормы или патологии. 2. Поведение запасания пищи обеспечивает максимально возможную стабильность кормовой базы группы и, особенно, прибылого потомства, тем самым повышая как уровень выживаемости последнего, так и уровень их формирования в пределах психологической нормы. 3. При выращивании в неволе животных, которые в дальнейшем должны быть реинтродуцированы, необходимо содержать их в среде, обогащенной крупными непрозрачными предметами, что имеет решающее значение для формирования максимального уровня (специфичного для вида) способности к рассудочной деятельности. В дальнейшем это обеспечит способность к многократному решению элементарных рассудочных задач, часто возникающих при охоте, и, исходя из этого, успешность самой охоты, а в конечном итоге – обеспечит высокий уровень выживаемости реинтродуцированных животных. 4. В процессе постнатального онтогенеза хищническое и охотничье поведение развивается поэтапно. Важно, что каждый предыдущий этап является основой для последующего. Полноценное развитие каждого из них возможно только в так называемый «период чувствительности», свойственный данному этапу.(См. в тексте) 5. Подготовка крупных хищных животных к реинтродукции должна начинаться на раннем этапе постнатального онтогенеза. Следует учитывать, что первая реакция на потенциальную жертву не связана с пищевой мотивацией. Эта связь генерализованно образуется только после установления хищником факта съедобности жертвы. 6. Только после того, как у молодого хищника установится ассоциация между потенциальной жертвой и пищей, возможен переход к обучению хищника охоте. 7. В процессе подготовки к реинтродукции следует учитывать то, что хищникам, обладающим высокой социальностью, необходимо предоставлять возможность групповой охоты. Это нужно не только для того, чтобы члены группы при нападении на крупную жертву научились взаимосогласованным действиям, которые способствуют успеху охоты, но и для развития навыков самой охоты. Все это формируется только путем накопления опыта методом проб и ошибок. 8. К моменту реинтродукции животные должны хорошо знать предназначенную для этого территорию. К началу ее освоения у них должна сформироваться «мысленная карта», обеспечивающая свободную ориентацию на территории. 9. К началу освоения территории, предназначенной для реинтродукции, необходимо начать выработку реакции избегания незнакомого человека и домашних животных. Это дает возможность предупредить конфликты между реинтродуцированными животными и местным населением. 10. Реакция избегания домашних животных и представителей местного населения должна вырабатываться при непосредственном участии последних. Это полностью меняет их отношение к восстанавливаемому виду, что имеет большое значение для успешной реинтродукции любого вида крупных хищных животных. 11. Реакция избегания домашних животных вырабатывается генерализованно. Ее следует выработать у хищника в отношении каждого вида, независимо от породы. Этот принцип выгоден и тем, что делает возможной выработку реакции избегания и в отношении диких животных, стоящих на грани исчезновения. А это даст возможность восстановить вид хищника без его отрицательного воздействия на исчезающие виды животных. 12. Дистанция бегства копытных от врага (человека и хищника) и интенсивность самой реакции избегания подвержены сильным модификациям в ходе онтогенеза, поэтому возможно их изменение во времени соответственно существующей ситуации. 13. В процессе восстановления вида хищных животных необходимо реинтродуцировать их небольшими группами, во избежание их отрицательного воздействия на численность жертвы. Интервалы между реинтродукциями должны быть не менее одного года, что будет достаточным для выработки поведенческого иммунитета к хищнику у видов жертвы. 14. Исходя из специфики высшей нервной деятельности, разработанный нами метод может быть применен к любому виду крупных хищных млекопитающих. Кроме того, метод освоения территории и выработки реакции избегания незнакомых людей применим и при реинтродукции копытных животных.
ПОСЛЕСЛОВИЕ
Как мы убедились, подготовка к реинтродукции в природу выращенных в неволе крупных млекопитающих и хищных, в частности, – проблема сложная и многоплановая. Тут существует ряд и других проблем. Как справедливо замечает Д. Ялден (Yalden, 1993), необходимо прежде всего аннулировать те факторы, которые вызвали исчезновение (или падение численности) этого вида. Далее автор подчеркивает важность содержания в неволе животных, способных размножаться. Вместе с тем он отмечает, что необходимо наличие «достаточного количества человеческих и финансовых ресурсов, чтобы программу реинтродукции довести до той стадии, когда шанс ее реализации будет довольно высок» (с. 299). К счастью, недостатка в человеческих, в том числе и интеллектуальных ресурсах, нет. Об этом свидетельствует та обширная и успешная работа, которая проводится в Европе и Америке. Другое дело – финансирование работ по охране и восстановлению исчезающих видов животных. Подобное могут позволить себе только экономически развитые страны. Экономически же слабые страны способны решить проблему охраны и восстановления исчезающих видов только с помощью финансовой поддержки Мирового Сообщества. Еще одна проблема, которая в конечном итоге может предопределить успех или провал любой программы реинтродукции крупных млекопитающих и, в частности, хищных – это отношение местного населения к реинтродуцируемому виду. Опыт показал, что никакая финансовая компенсация за нанесенный хищником урон не меняет отрицательного отношения человека к животному. В результате, сколько бы не было затрачено средств на компенсацию, животные все-таки гибнут от рук человека. Кроме всего перечисленного, как оказалось, существует еще одна не обсуждавшаяся ранее проблема – это проблема выращивания в неволе поведенческий полноценных животных, у которых наряду с этим максимально развита способность к рассудочной деятельности. Я надеюсь, что материал, приведенный в предлагаемой читателю книге, создаст основу для решения всех практических проблем, возникающих при содержании и подготовке к реинтродукции выращенных в неволе крупных хищных млекопитающих. Еще раз хочу особо подчеркнуть значение участия представителей местного населения в процессе выработки у животных реакции избегания незнакомых людей и домашних животных, т.к. только собственный опыт способен убедить людей в том, что хищник может стать неопасным для них самих и их собственности.
Литература
1. Анохин П.К. Рефлекс цели как объект физиологического анализа. «Журнал высшей нервной деятельности». 1962, т.126, вып.1, с.7-21. 2. Бадридзе Я.К. Запасание пищи волками. В сб.: Экологические основы охраны и рационального использования хищных млекопитающих. М., «Наука», 1979, с.83-84. 3. Бадридзе Я.К. О возможности функциональной трансформации реакции массажа лапками молочной железы во время сосания, на примере некоторых псовых. «Сообщения АН ГССР», 1982, т.107, №2, с.414-415. 4. Бадридзе Я.К. (а) О физиологическом механизме, запускающем поведение запасания пищи у собак. В сб.: Механизмы поведения. М.,»Наука», 1983а, с.186-189. 5. Бадридзе Я.К. (б) Поведение новорожденных волчат. В сб.: Механизмы поведения. М.,»Наука», 1983, с.134-135. 6. Бадридзе Я.К. Онтогенез пищевого поведения. В кн.: Волк. М.,»Наука», 1985, с.278-284. 7. Бадридзе Я.К. (а) Онтогенез пищевого поведения волка. Диссертация на соискание ученой степени кандидата биологических наук. М., МГУ, 1987. 139 с. 8. Бадридзе Я.К. (б). Пищевое поведение волка (вопросы онтогенеза). Тбилиси, «Мецниереба». 1987. 86 с. 9. Бадридзе Я.К. Реинтродукция в природу выращенных в неволе крупных хищных млекопитающих. Проблема и метод. Диссертация на соискание степени доктора биологических наук. М., МГУ, 1997. 147 с. 10. Бадридзе Я.К. Волк.I. Некоторые данные о формировании охотничьего поведения диких волков. Тбилиси, «Мецниереба», 1996. 17 с. 11. Бадридзе Я.К., Гуриелидзе З.В., Тодуа С.Г. О некоторых процессах в популяции благородного оленя, протекающих на фоне снижения численности волка. Сообщения АН Грузии. 1992, т.145, #3, с.607-610. 12. Бадридзе Я.К., Копалиани Н.Г., Гуриелидзе З.В., Тодуа С.Г., Бутхузи Л.Т. Волк. II. Формирование хищнического и охотничьего поведения у выросших в неволе волков. Тбилиси, «Мецниереба», 1996. 21 с. 13. Бакурадзе А.Н., Чхенкели С.А. (а). О некоторых механизмах формирования состояния насыщения. В кн.: Механизмы нейро-гуморальной регуляции вегетативных функций. М., «Наука», 1970, с.157-163. 14. Бакурадзе А.Н., Чхенкели С.А. (б). О механизмах нейро-гуморальной регуляции гипоталамуса. В сб.: Проблемы физиологии гипоталамуса. Выпуск 4, изд-во КГУ, 1970, с.3-10. 15. Барабаш-Никифоров И.И., Формозов А.Н. Териология. «Высшая школа», 1963. 394 с. 16. Бараташвили Т.К. Некоторые аспекты поведения волка в Грузии. В сб.:Прикладная этология. М., «Наука», 1983, т.3, с.195-197. 17. Бараташвили Т.К. Грузия и Армения. В кн.: Волк. М., «Наука», 1985, с.497-501. 18. Баскин Л.М. Поведение копытных животных. М., «Наука», 1976. 293 с. 19. Бериташвили И.С. Память позвоночных животных, ее характеристика и происхождение. Тбилиси, изд-во АН ГССР, 1968. 137 с. 20. Бериташвили И.С. Структура и функция коры большого мозга. М.,»Наука», 1969. 520 с. 21. Вырыпаев В.А. Об охотничьем поведении волка в Тянь-Шане. Зоологический журнал. 1980, т.59, вып.12, с.1874-18880. 22. Гращенков Н.И., Латаш Л.П. Об активном характере ориентировочной реакции. В сб.: Рефлексы головного мозга. Международная конференция, посвященная 100-летию выхода в свет одноименного труда И.М.Сеченова. М.,»Наука», 1965, с.263-275. 23. Дииник Н.Я. Кавказский олень. В кн.: Материалы к познанию фауны и флоры Российской Империи. М., 1904, вып.6. 24. Зворыкин Н.А. Волк и борьба с ним. М.-Л. КОИЗ, 1936. 120 с. 25. Игнатовская Н.Б. Отношение к животным как проблема нравственности. В кн.:Этическая мысль. М., Политиздат, 1988, с.220-237. 26. Козлов В.В. Волк. «Охота и охот. хоз-во». 1964, #9, с. 18-20. 27. Копалиани Н.Г., Бадридзе Я.К. Роль игры в формировании единоборства с жертвой у волков. Тбилиси, Изд-во АН Грузии, «Мецниереба», 1997. 19 с. 28. Корытин С.А. О поведение волчат. В сб.: Поведение охот.животных. Киров, 1981, с.87-94. 29. Корытин С.А., Бибиков Д.И. Охотничье поведение. В кн.: Волк. М.,»Наука», 1985, с.311-324. 30. Корытин С.А., Заболодских Ю.С. Видовые особенности пищевого поведения зверей семейства волчьих (Canidae). В сб.:Поведение охот.животных. Киров, 1981, сс.73-79. 31. Кочетков В.В. Наблюдения за охотничьим поведением волка в Средней Росии. В сб.: Поведение волка. М., изд-во АН СССР, 1980, с.60-75. 32. Красная Книга ГССР. Тбилиси, изд-во «Сабчота Сакартвело», 1982, 255 с. 33. Крушинский Л.В. Экстраполяционные рефлексы, как элементарная основа рассудочной деятельности у животных. «Доклады АН СССР», 1958, т.121, вып.4, с.762-765. 34. Крушинский Л.В. Формирование поведения животных в норме и патологии. М., изд-во МГУ, 1960. 264 с. 35. Крушинский Л.В. Есть ли разум у животных?» Природа», 1968, #9, с.75-78. 36. Крушинский Л.В. Биологические основы рассудочной деятельности. Эволюционные и физиологический аспекты поведения. М., изд-во МГУ, 1977. 269 с. 37. Крушинский Л.В., Дашевский Б.А., Сотская М.Н., Дудоладова Л.А., Мычко Е.Н., Шубкина А.В., Ермакова Е.В. Исследования элементарной рассудочной деятельности волка (Canis Lupus). Зоологический журнал. 1980, т.59, #6, с.915-924. 38. Крушинский Л.В., Молодкина Л.Н., Попова Н.П., Светухина В.М., Мац В.М. Экспресс-информационные ассоциации как основа некоторых сложных форм поведения животных. В сб.: Сложные формы поведения. М.-Л., «Наука», 1965, с. 58-63. 39. Крушинский Л.В., Мычко Е.Н., Сотская М.Н., Шубкина А.Н. Элементарная рассудочная деятельность и сложные формы поведения. В кн.: Волк. М., «Наука», 1985, с. 284-295. 40. Крушинский Л.В., Пажетнов В.С., Шубкина А.В. Изучение рассудочной деятельности бурых медведей. В сб.: Экология, морфология и охрана медведей в СССР. М., Главприрода МСХ СССР, 1981, с. 12-13. 41. Кудактин А.Н. Волк на Западном Кавказе. «Охота и охот. хоз-во», 1977, #9, с. 18-19. 42. Кудактин А.Н. (а). Пищевая специализация волка. «Охота и охот. хоз-во», 1978, #3, с.9-10. 43. Кудактин А.Н. (б). Об избирательности охоты волка на копытных в кавказском заповеднике. «Бюл. Моск. о-ва испытателей природы». Отд. биол. 1978, т.33, #3, с.19-28. 44. Кудактин А.Н. Охотничьи повадки волка на Западном Кавказе. «Бюл. Моск. о-ва испытателей природы». Отд.
|
||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 374; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.102 (0.013 с.) |