Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Значение техники искусственного выкармливания новорожденных хищных млекопитающих для формирования поведения взрослыхСодержание книги
Поиск на нашем сайте
Поведенческие реакции, сформированные к моменту рождения незрелорождающихся животных и способствующие их первому контакту с окружающей средой, довольно немногочисленны и, судя по литературным данным (Уждавини, 1958; Уждавини, Шепелова, 1966; Слоним, 1967, 1976; Хайнд, 1975; Бадридзе, 1982, 1983 б, 1985, 1987 а, б; Harlow & Zimmerman, 1959 и др.), довольно хорошо изучены. Они «как бы пригнаны к условиям существования сразу же после рождения» (Уждавини, Шепелова, 1966, с.106). О подготовленности реакций новорожденных к экологии вида писал и академик П.К.Анохин (1962). Опыты, проведенные в лаборатории А.Д.Слонима (1976), показали, что наиболее выражены, запрограммированные в эмбриональном периоде реакции на кормящую мать. Авторы установили, что можно не только наблюдать врожденные реакции на мать как на внешний фактор, но и моделировать эти внешние факторы. Подобное моделирование проводили и мы (Бадридзе, 1983 а, б; 1985, 1987 а, б). Такой подход способствует выявлению тех раздражителей, на которые возникают конкретные реакции новорожденного животного, что позволяет заполнить пробелы в изучении экологии неонатального периода тех видов, условия обитания которых не дают или ограничивают возможность исследований. Даже в таких значительных работах, как исследования Фокса (Fox, 1971) и Мича (Mech, 1970), касающихся поведения и экологии представителей семейства Canidae и, в частности, волка, нет каких-либо существенных данных об экологической адекватности поведения новорожденных животных. Это можно объяснить большой сложностью наблюдений в природе за контактом новорожденных животных (особенно рождающихся в логове) с матерью и остальной внешней средой, в ответ на воздействие которых развиваются строго определенные врожденные реакции. В лабораторных условиях Фокс (Fox, 1964, 1971) детально исследовал и описал неонатальные реакции Canidae и, в том числе, волка. К сожалению, ни в одной из упомянутых работ нет данных о том, какую роль играет полноценная реализация неонатальных реакций для формирования нормального поведения в постлактационном периоде. Поэтому в данной главе мы попытаемся показать значение полноценной реализации врожденных поведенческих элементов, связанных с процессом питания новорожденных хищных млекопитающих для дальнейшего формирования поведения в пределах нормы.
Материал и методика Эксперименты проводили на 96 новорожденных волчатах, 67 щенках собак (далее – щенки), 30 новорожденных котятах. 10 волчат, 15 щенков и 10 котят были взяты у матери в момент рождения. Возраст отнятия от матери 37 волчат и 15 щенков составлял до 40 часов, а 49 волчат и 37 щенков – 7-10 дней. Всех животных вскармливали с помощью бутылок с соской. Во время кормления 20 волчат и 15 щенков (I контрольная группа) на бутылку с соской надевали диск, диаметр которого составлял 200 мм (рис. 2А), чтобы у новорожденных была возможность упираться лапами во время сосания. Прокол в соске был таков, чтобы не позволять молоку вытекать без сосательных усилий. 10 волчат, 15 щенков и 10 котят (I экспериментальная группа) с момента рождения кормили из сосок без диска, тем самым лишая их возможности упираться во что-либо передними лапками. Остальных котят до окончания лактационного периода вскармливала мать. Во всех случаях у основания бутылки было проделано отверстие диаметром 1 мм для уравнивания давления во время сосания внутри бутылки с атмосферным. Значение этого в свое время было описано (например, Бадридзе, 1987 а, б).
Рисунок 2. Типы применяемых сосок (А, Б, В). 1. Диск; 2. Прокол; 3. Отверстие – 2мм; 4. Отверстия.
При вскармливании животных II экспериментальной группы (10 волчат и 10 щенков, отнятых у матери в возрасте 6-40 часов) также применяли два типа сосок: I тип – с проколом, который не позволял молоку вытекать без сосательных усилий, тем самым приближая процесс сосания к естественному (рис. 2Б) и II тип – с отверстием диаметром 2 мм (рис. 2В). При сосании соски с большим отверстием детенышам не приходилось прилагать значительных усилий, и желудок у них наполнялся после двух-четырех сосательных движений.
Рисунок 3. Приспособление, имитирующее тело матери. 1-резервуар с молоком; 2-трубки, проводящие молоко; 3-резиновый баллон с водой 36-38оС; 4-кроличья шкурка; 5-искусственные сосцы.
Остальных животных (II контрольная группа, 56 волчат и 27 щенков) вскармливали с помощью сконструированной нами установки, имитирующей тело матери. Установка представляла собой покрытый кроличьими шкурками изогнутый дугой резиновый баллон (рис.3). Сосцы прикреплялись к баллону со стороны меньшего радиуса. Они соединялись мягкими пластиковыми трубками, пронизывающими баллон. В полость баллона заливалась теплая вода (36-38 градусов), которая нагревала находящееся в трубках молоко. Пастеризованное молоко для вскармливания щенков и волчат приготавливали из молочного порошка, имитируя концентрацию собачьего молока (Барабаш – Никифоров, Формозов, 1963). В результате наблюдения за животными в условиях искусственного вскармливания нами были изучены следующие вопросы: 1. Влияние полноценной реализации «реакции массажа лапками молочной железы» во время сосания на формирование психически уравновешенного взрослого животного. 2. Влияние скорости поступления молока во время сосания в первую неделю жизни на дальнейшее формирование порога сенсорного насыщения. Полученные результаты и их обсуждение В свое время мы показали, что «реакция массажа лапками молочной железы» во время сосания в постлактационный период трансформируется в манипуляторную активность при разделке пищи (Бадридзе, 1982, 1983 б, 1985, 1987 а, б. Рис. 4 А, Б).
Рисунок 4. Функциональная трансформация реакции массажа лапками молочной железы во время сосания. А – лактационный период; Б – постлактационный период.
«Реакция массажа лапками молочной железы» во время сосания представляет собой результат последовательного сокращения разгибательных и сгибательных мышц передних конечностей. В наших экспериментах, когда животных лишали возможности упора лапками во что-нибудь (I экспериментальная группа – 15 щенков, 10 волчат и 10 котят), у них появлялось тоническое напряжение передних конечностей. Это выражалось в растопыривании пальцев и развитии одновременного напряжения сгибательных и разгибательных мышц. Подобное состояние продолжалось до окончания акта сосания. Те новорожденные (I и II контрольные группы), у которых постоянно была возможность опираться на что-либо передними лапками (бутылочка с диском, либо прибор, имитирующий тело матери), осуществлялась нормальная «реакция массажа лапками», т.е. последовательное сокращение реципрокных мышц передних конечностей. В возрасте, когда хищники переходят на полный мясной рацион (40-50 дней), животным начали давать по крупному куску мяса. При первом же его предъявлении все животные контрольных групп начинали его разделывать причем с первого же момента все манипуляционные движения лап были полностью координированы. Иная картина наблюдалась в I экспериментальной группе. Здесь для формирования координированной манипуляторной активности требовалось 3-4 дня. У этих животных заметно проявлялось т.н. «переступание лапами на месте». Параллельно с этим, животные время от времени растопыривали пальцы. Со временем тик ослабевал, но в малозаметной форме сохранялся до конца эксперимента. Кроме того, у всех животных в течение всего периода наблюдений была заметна эмоциональная неуравновешенность, что проявлялось в сравнительно низком пороге оборонительного поведения и частых фрустрациях. Эти индивиды часто создавали в группе (с т.н. нормальными) конфликтные ситуации. Соотношение животных контрольной и экспериментальной групп, провоцирующих конфликт, составляло 1/1, 25. Волчата и щенки контрольных групп развивались эмоционально нормальными и способны были создать группу с социальными отношениями в пределах нормы. До обсуждения возможных причин подобного различия считаем нужным напомнить данные, приведенные Г.Шепердом (1987), согласно которым импульсация от мышечных и суставных рецепторов поступает через таламус в соответствующую область коры головного мозга. Общеизвестно, что у незрелорождающихся млекопитающих формирование мозговых систем продолжается после рождения и, судя по всему, оптимальный уровень центростремительной импульсации имеет большое значение для их нормального формирования и функционирования. Во время сосания у животных I экспериментальной группы постоянно развивалось тоническое напряжение мышц передних конечностей, что должно было определять центростремительную гиперимпульсацию к соответствующим мозговым структурам. Из сказанного можно предположить, что в период формирования мозговых систем подобная гиперимпульсация может создавать очаги с постоянно высоким уровнем возбуждения, что, в свою очередь, влияет на уровень эмоциональной уравновешенности животного. Из приведенного материала следует, что животные, выращенные при подобных условиях питания в неонатальном периоде, не могут быть пригодными для дальнейшей их реинтродукции. Сравнивая различия в уровне эмоциональной уравновешенности между животными I экспериментальной и контрольными группами, можно заключить, что из-за невозможности нормальной реализации «реакции массажа лапками молочной железы во время сосания» (I экспериментальная группа), развивается одновременное тоническое напряжение всех групп мышц передних конечностей, вызывает возникновение в мозгу очагов постоянно высокого возбуждения. Судя по всему, это и стало причиной эмоциональной неуравновешенности животных этой группы. Животные же контрольных групп имели возможность реализации данной реакции, что, по видимому, и определило нормальное формирование их эмоциональной возбудимости, т.к. у них в мозгу не возникают очаги с постоянно высоким возбуждением. Как было сказано ранее, животных II экспериментальной группы кормили с помощью сосок с отверстием диаметра 2 мм. У волчат и щенков, вскармливаемых из такой соски, желудки наполнялись после нескольких сосательных движений, в течение 0, 5 минуты (60-80 мл). Во время подобных экспериментов все животные, несмотря на быстрое наполнение желудка, продолжали проявлять сосательную активность. В свое время нами было показано, что у новорожденных щенят и волчат пищевая мотивация возникает к концу первой недели жизни (Бадридзе, 1987 а, б), а до этого процесс приема пищи осуществляется за счет потребности в реализации сосательной активности. Животные этой группы продолжали принятие чрезмерного количества пищи и после формирования пищевой мотивационной системы. Стремление к перееданию сохраняется в течение всей жизни, что создает постоянные внутригрупповые конфликты на почве конкуренции за пищу. В свое время было показано (Анохин, 1962; Бакурадзе, Чхенкели, 1970 а, б; Бадридзе, 1987 а, б и др.), что включение центральных механизмов головного мозга, тормозящих акт питания, происходит за счет сенсорного насыщения, т.е. центростремительной импульсации, возникающей при растяжении стенок желудка. Анализируя полученный нами материал, можно заключить, что чрезмерный прием пищи в раннем неонатальном периоде может увеличить объем желудка и, следовательно, порог сенсорного (первичного) насыщения, что и определяет развитие относительной гиперфагии. Такие животные активно конкурируют из-за пищи до наступления вторичного насыщения, чем постоянно создают конфликтную ситуацию во время еды. Вместе с тем у них, как правило, не развивается поведение запасания пищи, значение которого будет обсуждаться в следующей главе. Из всего вышесказанного следует, что для нормального развития новорожденных животных, отлученных от матери, решающее значение имеют т.н. «технические» условия при приеме пищи, которые максимально должны быть приближены к естественным. Животные, выращенные в иных условиях, не могут быть пригодны для реинтродукции, т.к. не способны поддерживать в группе полноценные социальные взаимоотношения. Из полученного нами материала можно сделать следующий основной вывод – при искусственном вскармливании хищных млекопитающих для нормального формирования поведения особое значение имеет неонатальный период, который в значительной мере определяет дальнейшее развитие поведения в пределах нормы или патологии. В частности: 1. для формирования оптимального уровня эмоциональной возбудимости большое значение имеет возможность полноценной реализации реакции «массажа лапками молочной железы во время сосания»; 2. для исключения высокого порога сенсорного насыщения и, исходя из этого, высокого уровня конкурентных ситуаций в группе во время питания, необходимо условия выкармливания животных максимально приблизить к естественному процессу сосания.
Глава 1.2. Поведение запасания пищи
Запасание корма является одним из врожденных видов пищевого поведения, широко распространенного среди животных (Свириденко, 1957; Слоним, 1967; Хайнд, 1975 и др.) Из хищных млекопитающих пищей запасаются собаки, волки, лисицы и др. (Мертц, 1953; Кудактин, 1977; Поярков, 1980; Корытин, 1981; Jeselnik, Brisbin, 1980 и др.) Наиболее полные из имеющихся в литературе сведений по этому вопросу можно найти у Мича (Mech, 1970), однако и их явно недостаточно для исчерпывающего понимания феномена запасания пищи волками. Мич также подчеркивает, что о запасании волками пищи и о том, как это происходит, известно очень мало. Упоминания об этом поведении волка содержатся и в работах Н.А. Зворыкина (1936) и Б.Т.Семенова (1979). В свое время мы изучали закономерность формирования поведения волков при запасании пищи, зависимость способа транспортировки запасаемой пищи от социального статуса особи и специфику памяти на местонахождение кладовых (Бадридзе, 1979, 1987 а, б; Бадридзе и соавт., 1997). К сожалению, мы не встречали в литературных источниках данных, касающихся значения данного вида поведения как для индивида, так и для всей группы в целом и, исходя из этого, для всей популяции. Неизвестно также, может ли существовать естественная патология в формировании данного поведения. Мы имеем в виду естественную патологию, так как причины, из-за которых не формируется поведение запасания пищи при искусственном выкармливании новорожденных животных, уже обсуждались в главе 1.1. Поэтому в данной главе будут освещены вопросы, касающиеся значения поведения запасания для пищевого благополучия индивида, группы и, исходя их этого, для популяции в целом. Изучение данного вопроса должно иметь большое значение для отбора животных по признаку сформированности этого поведения к моменту реинтродукции в природу.
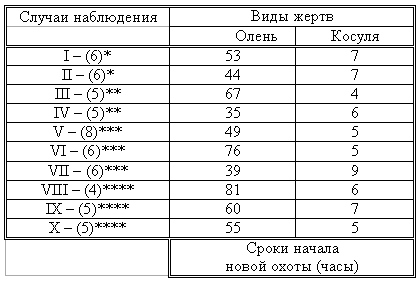
Материал и методика Наблюдения проводили на 78 ручных волках, выращенных в неволе с первых дней жизни до 5-месячного возраста, а на 39 из них – до полового созревания. Помимо этого приведен материал наблюдений за 4 семьями диких волков, приученных к моему присутствию (820 часов наблюдений). Забегая вперед, скажем, что в этой же главе обсудим материал о поведении запасания, полученный на реинтродуцированных нами четырех группах волков. Полученные результаты и их обсуждение Наблюдения за ручными волками показали, что поведение запасания сформировалось только у 68 волчат. Следует отметить, что из 10 животных, у которых данный вид поведения не сформировался, 8 входили в экспериментальную группу, и причиной несформированности поведения запасания в этом случае была, по-видимому, невозможность осуществления «массажа лапками молочной железы» во время сосания. Два волчонка были представителями контрольной группы, порог сенсорного насыщения в которой сформировался в пределах нормы. Исходя из сказанного, причины несформированности данного поведения у этих животных надо искать во врожденном дефекте, природа которого нам пока непонятна. Мы учитываем этот факт для дальнейшего обсуждения вопроса значимости поведения запасания пищи у волков. В свое время мы установили (Бадридзе, 1987 а, б), что при достаточном количестве добычи, волки после еды 3-4 раза прячут пищу. Надо отметить, что каждый волк запасает от 0, 5 до 3, а иногда до 6 кг мяса (последнее зарегистрировано только 4 раза). Средний же вес запаса составлял около 2 кг без учета указанного максимального количества (всего проведено замеров веса содержимого 560 кладовых). Было определено среднее количество особей в группе дикоживущих волков, которое близко к 6 (проведены учеты 83 стай волков; учитывались только взрослые и переярки, т.к. количество запасаемой волчатами пищи менялось в зависимости от их возраста). Если принять число 6 как среднее количество индивидов в стае, то при достаточных размерах жертвы можно представить приблизительное количество запасаемой пищи, которое составит около 36 кг при 3-кратной реализации поведения запасания каждым индивидом. Наблюдения за дикими и реинтродуцированными нами волками показали, что из остатков их трапезы другие плотоядные подбирают, в первую очередь, недоеденные куски, оставшиеся на поверхности. Нанесенные ими потери невелики. В интервалах между охотой волки часто наведываются к запасенной пище. Наши наблюдения в этом плане вполне соответствуют приведенным в литературе данным, которые убедительно показывают, что волки в неблагоприятные для охоты периоды используют кладовые, расположенные в районе добывания пищи (например: Поярков, 1980). Необходимо отметить, что, когда прибылым исполняется 3 месяца, родители в интервале между удачной охотой часто приводят волчат к месту предыдущей трапезы, где они остаются в течение нескольких дней и полностью утилизируют все запасы. Подобное мы постоянно отмечали, наблюдая за дикими и реинтродуцированными нами волками. Даже если учесть, что 3-4-месячные волчата за счет мышкования и ловли насекомых могут на 50% обеспечить себя пищей (о чем пойдет речь во втором разделе), то можно представить значение запасания пищи взрослыми индивидами как фактора, обеспечивающего волчатам максимально возможную стабильность кормовой базы. Естественно возникает вопрос – что же происходит, когда размер добычи недостаточен для запасания? Наблюдения показали, что, если добычи достаточно для одноразового полного насыщения стаи, но недостаточно для запасания, количество попыток охоты возрастает. Однако это вовсе не гарантирует сокращение интервала между удачными охотами. В тех случаях, когда размер добычи позволяет волкам полностью насытиться, но ее недостаточно для запасания, новая охота начинается спустя 5-7 часов. В случае крупной добычи, когда поведение запасания реализуется в достаточной мере, охота начинается через 2-4 суток. Эти данные приведены в таблице №2. Необходимо отметить, что у взрослых и переярков количество актов запасания снижается, как только прибылым исполняется 7 месяцев, после чего последние сами интенсивно запасают пищу. А после того, как прибылым исполняется 9 месяцев и они начинают участвовать в охоте взрослых, старшие запасают не более 1 раза.
Таблица 2. Зависимость времени начала новой охоты, от размера добытой жертвы. В скобках обозначено количество членов семьи, включая переярков.
Таким образом, уровень поведения запасания пищи максимально возрастает в период, когда необходимо обеспечить стабильную кормовую базу для прибылых. В основном это происходит до выхода прибылых из ювенильного возраста. Об интенсивном запасании пищи волками в период выкармливания потомства пишет В.Козлов (1964). Полученный нами материал свидетельствует о том, что поведение запасания пищи представляет собой форму поведения, которая в определенной мере способна обеспечить прибылое потомство стабильной кормовой базой. Поскольку взрослым волкам приходится обеспечивать пищей довольно многочисленное потомство, которое с 3-месячного возраста потребляет почти столько же мяса, сколько взрослые волки, не участвуя при этом в охоте, этот период, на наш взгляд, следует отнести к наиболее трудным в жизни группы. Взвешивая реинтродуцированных нами волков после полного насыщения добытой ими пищи и неоднократной реализации поведения запасания, мы определили, что чистый вес съеденного мяса составлял в среднем 3, 5 кг. Если учесть, что каждый волк запасает от 6 до 8 кг, то присутствие в группе одного не запасающего индивида может нанести значительный урон в обеспечении прибылых кормом. Из этого следует, что у животных, отобранных для дальнейшей реинтродукции в природу, поведение запасания пищи должно быть полностью сформировано. Исследовав принципы формирования агрессивности волчат в ювенильном периоде при их 50%-й пищевой депривации, мы пришли к заключению, что постоянное недоедание меняет приоритеты психологического комфорта (см. Овсянников, Бадридзе, 1989), вследствие чего агрессивное поведение не ритуализируется (Бадридзе, 1997). Вряд ли стоит обсуждать значение ритуализации агрессивного поведения. Итак, феномен запасания пищи у волков представляет собой генетически детерминированный поведенческий акт. Он становится интенсивным во время выкармливания прибылого потомства, тем самым обеспечивая максимально возможную стабильность кормовой базы для волчат в ювенильном периоде. В тех же случаях, когда размер добычи не позволяет делать запасы, интервал между насыщением и охотой значительно сокращается. Это повышает уровень как выживаемости прибылого потомства, так и их формирования в пределах психологической нормы. Резюмируя приведенный материал, можно сделать следующие выводы: поведение запасания пищи приобретает особое значение в период выкармливания прибылого потомства; невозможность реализации поведения запасания стимулирует сокращение интервалов между попытками охоты; 1. наличие в группе даже одной особи, у которой не сформировано поведение запасания пищи, отрицательно сказывается на стабильности кормовой базы волчат; они постоянно недоедают, вследствие чего растет их агрессивность на почве пищевой конкуренции; 2. особи, у которых не формируется поведение запасания пищи, не должны отбираться для реинтродукции в природу.
Глава 1.3
|
||||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 195; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.224.54.118 (0.009 с.) |