Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Химизм реакции цикла КальвинаСодержание книги
Поиск на нашем сайте
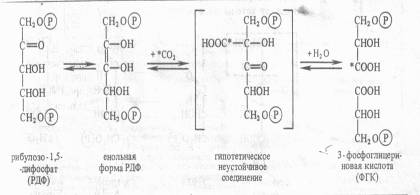
Этот способ ассимиляции С02, присущий всем растениям. Фотосинтезирующие водоросли помещали в среду, содержащую 14С02, на разные промежутки времени, затем клетки быстро фиксировали, экстрагировали из них спирторастворимые вещества и определяли содержание 14С в различных соединениях после их разделения с помощью хроматографии. Оказалось, что после экспозиции в течение 1 мин 14С включался в С3 —С7-сахара и фосфосахара, в органические кислоты (яблочную, щавелевоуксусную, ФЕП), в аминокислоты (аланин, аспарагиновую кислоту). Если же время экспозиции было сокращено до 0,1—2 с, то большая часть метки обнаруживалась в фосфоглицериновой кислоте, в ее карбоксильной группе. Следовательно, 3-фосфоглицериновая кислота (ФГК) является первичным продуктом фотосинтеза. Природа первичного акцептора С02. Сначала предположили, что таким акцептором является какое-либо двухуглеродное соединение. Однако введение в инкубационную среду винилфосфата, фосфогликольальдегида и других веществ с С2 не приводило к увеличению содержания радиоактивной метки из С02 в ФГК. Тогда схему опыта видоизменили следующим образом. Водоросли экспонировали на свету при высокой (1 %-ной) концентрации С02, затем резко снижали его концентрацию до 0,003%. Расчет был на то, что в условиях дефицита С02 быстро накопится именно то соединение, которое служит акцептором С02. С помощью двумерной хроматографии удалось установить, что при отсутствии возможности карбоксилирования в клетках кратковременно возрастает концентрация рибулозо-1,5-дифосфата (рибулозо-1,5-бисфосфата). Отсюда возникло предположение, что первичная фиксация С02 происходит следующим образом: С5 + С02-> С6-► 2С3. Для проверки этой гипотезы в бесклеточный экстракт из листьев шпината или клеток хлореллы вносили меченный по 32Р рибулозо-1,5-дифосфат. На свету в экстракте появлялась радиоактивная ФГК. На основании полученных данных процесс первичной фиксации С02 можно записать следующим образом:
Эта реакция катализируется рибулозодифосфаткарбоксилазой (рибулозобисфосфаткарбоксилазой; другие названия — белок фракции I, карбоксидисмутаза). Максимум активности фермент имеет при рН 7,8 — 8,0 и нуждается в присутствии ионов Mg2 +. Дальнейшая работа в лаборатории Кальвина и в других лабораториях привела к расшифровке всех последующих реакций С3-пути фотосинтеза, который получил название цикла Кальвина. Этот цикл, весьма напоминающий обращенный пентозофосфатный путь дыхания, состоит из трех этапов: карбоксилирования, восстановления и регенерации. 1. Карбоксилирование. Молекулы рибулозо-5-фосфата фосфорилируются с участием АТР и фосфорибулозокиназы, в результате чего образуются молекулы рибулозо-1,5-дифосфата, к которым в свою очередь присоединяется С02 с помощью рибулозодифосфаткарбоксилазы. Полученный продукт расщепляется на две триозы: 2 молекулы 3-фосфо-глицериновой кислоты (3-ФГК). 2. Фаза восстановления. 3-ФГК восстанавливается до 3-фосфоглицеринового альдегида (3-ФГА) в два этапа. Сначала происходит фосфорилирование 3-ФГК при участии АТФ и фосфоглицераткиназы до 1,3-дифосфоглицериновой кислоты, а затем восстановление 1,3-ФГК с помощью НАДФH и дегидрогеназы фосфоглицеринового альдегида. 3. Фаза регенерации первичного акцептора диоксида углерода и синтеза конечного продукта фотосинтеза. В результате описанных выше реакций при фиксации трех молекул С02 и образовании шести молекул восстановленных 3-фосфотриоз пять из них используются затем для регенерации рибулозо-5-фосфата, а один — для синтеза глюкозы. 3-ФГА под действием триозофосфатизомеразы изомеризуется в фосфодиоксиацетон. При участии альдолазы 3-ФГА и фосфодиоксиацетон конденсируются с образованием фруктозо-1,6-дифосфата, у которого отщепляется один фосфат с помощью фруктозо-1,6-дифосфатазы. В дальнейших реакциях, связанных с регенерацией первичного акцептора С02, последовательно принимают участие транскетолаза и альдолаза. Транскетолаза катализирует перенос содержащего два углерода гликолевого альдегида от кетозы на адьдозу:
Альдолаза затем осуществляет перенос трехуглеродного остатка фосфодиоксиацетона на альдозу, в данном случае эритрозо-4-фосфат, в результате чего синтезируется седо-гептулозо-1,7-дифосфат. Последний дефосфорилируется и под действием транскетолазы из него и 3-ФГА образуются ксилулозо-5-фосфат и рибозо-5-фосфат. Две молекулы ксилулозо-5-фосфата при участии рибулозофосфатэпимеразы и одна молекула рибозо-5-фосфата с участием рибозофосфатизомеразы превращаются в три молекулы рибулозо-5-фосфата, с которого начинается новый цикл фиксации С02. Из оставшейся неиспользованной 6-ой молекулы 3-ФГА под действием альдолазы синтезируется (при повторении цикла) молекула фруктозо-1,6-дифосфата, из которой могут образовываться глюкоза, сахароза или крахмал: Таким образом, для синтеза одной молекулы глюкозы в цикле Кальвина необходимы 12 NADPH и 18 АТР, которые поставляются в результате фотохимических реакций фотосинтеза. 11. Химизм реакций ассимиляции С₄ растений. Цикл Хетча-Слека-Карпилова. САМ-метаболизм органических кислот В работах Л. А. Незговоровой (1956—1957 гг.), было установлено, что при коротких экспозициях листьев кукурузы на свету 14С из 14С02 обнаруживается в аспарагиновой кислоте. В дальнейших исследованиях как советских, так и зарубежных специалистов эти представления были развиты, что привело к открытию C4-nymu углерода в фотосинтезе. Так, в 1960 г. Ю. С. Карпилов, а в 1963 г. И. А. Тарчевский и Ю. С. Карпилов представили данные о раннем образовании яблочной кислоты в листьях кукурузы. Г. П. Корчак и др. впервые показали, что дикарбоновые кислоты (яблочная и аспарагиновая) являются первичными продуктами фиксации С02 у сахарного тростника. Затем эти соединения через 3-ФГК и гексозофосфаты превращаются в сахара. Как новый тип фиксации С02, принципиально отличающийся от цикла Кальвина, этот цикл впервые описали австралийские ученые М. Д. Хетч и, К. Р. Слэк (1966). К группе растений с С4-путем фотосинтеза относятся сахарный тростник, кукуруза, сорго и др. Листья этих растений содержат два разных типа хлоропластов: хлоропласты обычного вида — в клетках мезофилла и большое количество крупных хлоропластов, часто не имеющих гран, — в клетках, окружающих проводящие пучки (обкладка). С02, диффундирующий в лист через устьица, попадает в цитоплазму клеток мезофилла, где при участии ФЕП-карбоксилазы вступает в реакцию с ФЕП, образуя щавелевоуксусную кислоту (оксалоацетат). Затем уже в хлоропластах оксалоацетат восстанавливается до яблочной кислоты (малата) за счет NADPH, образующего в ходе световой фазы фотосинтеза. Щавелевоуксусная кислота в присутствии NH4+ может превращаться также в аспартат. Затем малат (или аспартат) переносится в хлоропласты клетки обкладки сосудистого пучка, где он декарбоксилируется малик-энзимом (малатдегидрогеназой декарбоксилирующей) до пирувата и С02. В хлоропластах обкладки отсутствуют граны, а следовательно, слабо представлена ФС II, необходимая для нециклического транспорта электронов, однако в них в изобилии накапливается крахмал. Объясняется это тем, что в хлоропластах обкладки используется поставляемый малик-энзимом НAДФH, а также тот С02, который образовался при окислительном декарбоксилировании малата (или аспартата). В этих хлоропластах в процессе циклического фотофосфорилирования синтезируется большое количество АТР и фиксация С02 осуществляется по типу цикла Кальвина. У некоторых растений с С4-путем фотосинтеза (амарант, лебеда) яблочная кислота декарбоксилируется в митохондриях клеток обкладки с восстановлением NAD. Возникающий при расщеплении малата в хлоропластах клеток обкладки пируват перемещается назад в хлоропласты клеток мезофилла, где может снова превращаться в первичный акцептор С02 — ФЕП. Такая компартментация процессов позволяет растениям с С4-путем осуществлять фотосинтез даже при закрытых устьицах, так как хлоропласты клеток обкладки используют малат (аспартат), образовавшийся ранее, как донор С02. С4-растения могут также использовать С02, возникающий при фотодыхании. Закрывание устьичных отверстий в наиболее жаркое время дня сокращает потери воды за счет испарения (транспирации). Не удивительно поэтому, что к С4-растениям относятся многие виды засушливой тропической зоны. Растения с С4-путем фотосинтеза, как правило, устойчивы к засолению. Эффективность использования воды, т. е. отношение массы ассимилированного С02 к массе воды, израсходованной при транспирации, у С4-растений зачастую вдвое выше, чем у С3-растений. Таким образом, С4-растения имеют преимущества перед С3-растениями в засушливых местах обитания благодаря высокой интенсивности фотосинтеза даже при закрытых устьицах. Фиксация С02 с участием ФЕП и образование малата (аспартата) служит как бы насосом для поставки С02 в хлоропласты обкладки, функционирующие по С3-пути.
|
||||
|
|
Последнее изменение этой страницы: 2016-04-23; просмотров: 937; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.224.60.132 (0.01 с.) |