Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Эта работа была выполнена в рамках Государственной программы Геологического института Российской академии наук, Москва, Россия.Содержание книги
Поиск на нашем сайте
Обсуждение Изучена принадлежность хозяев челюстей головоногих моллюсков В мергелях Дивьинской свиты присутствуют внутренние формы раковин наутилоидов и аммоноидов, а также челюсти головоногих моллюсков. Наутилоиды представлены тремя отрядами: Oncocerida, Pseudorthocerida и Nautilida (Наугольных 2016, 2018) (рис. 5). Онкоцериды рода Scyphoceras Руженцева и Шиманского из семейства Scyphoceratidae (подотряд Rutocerina) являются самыми редкими головоногими моллюсками в этих отложениях (рис. 5А). Эти необычные киртоконические наутилоиды с широкой объемной камерой тела и небольшим фрагмоконом ранее рассматривались как представители отряда Nautilida (Руженцев и Шиманский, 1954). Однако многие исследователи в настоящее время рассматривают Rutoceratina как часть Oncocerida (например, Manda & Turek 2011), следовательно, Scyphoceras относится к этому отряду. Учитывая редкость Scyphoceras в Дивьинской формации и тот факт, что челюсти никогда не были найдены в ассоциации с онкоцеридами (Мироненко 2021), маловероятно, что описанные здесь челюсти принадлежат к этому роду. Свернутые спиралью наутилоиды (Metacoceras artiense Kruglov) также редки в дивьинской формации (рис. 5Б), и крайне маловероятно, что изученные челюсти принадлежали им из-за их редкости. Псевдортоцериды с прямым панцирем (Dolorthoceras siphocentrale (Кротов), Рис. 5. Головоногие моллюски из формации деления. А, онкоцеридные сцифоцерасы. B, наутилид Metacoceras artiense. C, pseudorthocerida Dolorthoceras sp. D, гониатитидный аммоноид Uraloceras sp. с нижней челюстью (образец K-R/1). E, увеличенное изображение нижней челюсти из D. Столбики шкалы: 1 см. Dolorthoceras sp.) встречаются чаще (рис. 5С); однако многое остается неясным о челюстях этой группы головоногих моллюсков. На сегодняшний день описано очень мало находок их челюстей. Поздний миссисипский гордониконус beargulchensis Maps, Weller & Doguzhaeva с сохранившимся челюстным аппаратом, который первоначально был описан как ортоконический наутилоид (Landman & Davis, 1988), в настоящее время считается представителем Coleoidea (Maps et al., 2010; Klug et al. 2019), поэтому его челюсти нельзя считать примером наутилоидных мандибул. Челюсти каменноугольной псевдортоцериды Reticycloceras Gordon также упоминались Мейпсом (1987), тем не менее фотографий, приведенных в этой публикации, недостаточно, чтобы понять форму и размер этих челюстей. Несмотря на это, Мэйпс (1987) процитировал устное сообщение Р.С. Кокса, касающееся головоногих моллюсков, собранных в миссисипских (каменноугольных) отложениях центральной Монтаны (США), в котором находки челюстей ортоконических наутилоидов гораздо реже, чем челюстей аммоноидов из тех же слоев и местностей.
Следовательно, наиболее вероятно, что пермские челюсти из Красноуфимского района принадлежат аммоноидам, поскольку раковины этих головоногих моллюсков гораздо многочисленнее в слоях дивьинской свиты, чем раковины наутилоидов, а размер и форма челюстей полностью совпадают с размерами и формой нижних челюстей ранее описанных аммоноидов. Скорее всего, челюсти принадлежат аммоноидам рода Uraloceras Ruzhencev (Paragastrioceratidae, Goniatitida), раковины которых наиболее многочисленны в Дивьинской формации. Это также подтверждается расположением одной из челюстей вблизи внутренней формы раковины Uraloceras (образец K-R/1, см. рис. 5D,E). Поскольку окаменелости в этих мергелях редки и большинство из них найдено на большом расстоянии друг от друга, такая близость вряд ли случайна. Принадлежность этих челюстей к гониатитидным аммоноидам также подтверждается их сходством с нижними челюстями миссисипского гониатитида Cravenoceras Bisat, которые также имеют большие внутренние пластинки (Kruta et al., 2014). Некоторые из образцов (рис. 3A,D) напоминают челюсти головоногих моллюсков верхнего мела, недавно описанные как возможные нижние челюсти колеоидных вампироподов (Klug et al., 2020). Однако, во-первых, о находках таких колеоидов в артинских отложениях никогда не сообщалось, а во-вторых, трапециевидная форма центральной платформы внутренних пластинок этих челюстей полностью идентична форме образцов K-R/2 и K-R/13 (рис. 3C, 3B соответственно), которые, несомненно, являются аммоноидными анаптихами. Следовательно, наиболее вероятно, что форма образцов K-R/7 и K-R/6 обусловлена их почти вертикальным расположением в осадке, тогда как они относятся к тому же типу, что и образцы K-R/2 и K-R/13. Следовательно, мы предполагаем, что все эти нижние челюсти являются аммоноидными анаптихами.
Интерпретация образцов верхних челюстей из Дивьинской свиты несколько сложнее. Они похожи на колеоидные челюсти, но в отсутствие находок колеоидных щитков или гладиусов, упомянутых выше, у нас нет оснований предполагать присутствие этих головоногих моллюсков в бассейне Приуралья-Артинского моря. Верхние челюсти пермских аммоноидов, насколько нам известно, еще не описаны. Вообще, существует интересный парадокс, который заключается в том, что нижние челюсти аммоноидов в ископаемом состоянии, как правило, значительно превосходят по численности верхние челюсти. Находки на верхних челюстях встречаются гораздо реже. Согласно Tanabe et al. (2015), находки челюстных аппаратов in situ известны из 109 родов аммоноидов, в то время как верхние челюсти известны только из 41 рода. Это хорошо отражено в истории изучения челюстей аммоноидов. В то время как аптихи и анаптихи (оба являются нижними челюстями) аммоноидов известны с 1702 года (см. исторический обзор в Trauth 1927), первая верхняя челюсть аммоноида была описана только полтора столетия спустя, в 1864 году (Meek & Hayden 1864). После долгого столетнего перерыва верхние челюсти были описаны во второй раз в 1967 году (Closs 1967; Lehmann 1967). К счастью, на сегодняшний день описано несколько образцов каменноугольных (Closs 1967; Mapes 1987; Bandel 1988; Догужаева и др. 1997) и один экземпляр верхнедевонских (Klug et al. 2016) аммоноидных верхних челюстей. Наши находки заметно отличаются от девонской верхней челюсти клаймениидного аммоноида Postclymenia, которая, как ни странно, напоминает верхние челюсти мезозойских аммонитов. Однако пермские образцы очень похожи на верхние челюсти аммоноидов гониатитид каменноугольного периода (Догужаева и др., 1997). Следовательно, мы предполагаем, что описанные здесь верхние челюсти, а также нижние челюсти принадлежат к гониатитидным аммоноидам рода Uraloceras.
Челюстной аппарат Uraloceras и вопрос о существовании палеозойских аптичей Согласно классификации типов челюстей аммоноидов (см. Tanabe et al., 2015), челюсти пермских Uraloceras следует отнести к нормальному типу, так же как и челюсти аммоноидов каменноугольного периода, из-за формы верхней челюсти. В то же время челюсти Uraloceras отличаются от челюстей позднекаменноугольных гониатитидных аммоноидов. Как верхняя, так и нижняя челюсти этих более древних гониатитид обычно имеют скульптуру, представленную тонкими ребрами и линиями роста (Догужаева и др., 1997; Догужаева 1999), в то время как пермские челюсти либо гладкие, либо покрыты редкими волнистыми ребрами. Таким образом, челюсти пермских гониатитид из дивьинской свиты больше похожи на челюсти аммоноидов позднего триаса типа anaptychus, чем на челюсти их предков каменноугольного периода. Ранее Карпинский (1890, 1891) описал нижнюю челюсть аммоноида из артинских отложений в окрестностях Красноуфимска, интерпретировав ее как aptychus. Аптихи - это парные симметричные элементы нижних челюстей типа аптихуса, которые, согласно современным представлениям, возникли в тоарском периоде (ранняя юра). Нижние челюсти типа aptychus разделены гибкой связкой (симфизом) по центральной линии, которая часто распадалась после смерти моллюска, и симметричные половины челюсти (аптихи) были погребены отдельно. Анаптичи, с другой стороны, представляют собой монолитные нижние челюсти, которые не разделены на легко отделяемые половины. Единственный экземпляр, изображенный Карпинским, действительно напоминает аптихуса. Однако мы предполагаем, что определение артинского экземпляра как aptychus было ошибочным. Судя по его схематическому изображению (Карпинский, 1890, рис. 56; Карпинский, 1891, рис. 15) этот образец почти идентичен описанному здесь образцу K-R/12 (рис. 2F), который представляет собой отпечаток правой половины анаптиха. Судя как по этому образцу, так и по другому подобному образцу (рис. 2E), Uraloceras anaptychi иногда были погребены в боковом положении, сложенные вдоль средней линии. Возможно, что срединная борозда, которая видна на одном образце (рис. 2D) могло бы облегчить такое сворачивание. Несмотря на то, что эта борозда может отражать появление более или менее подвижного симфиза между двумя половинами нижней челюсти, что впоследствии способствовало появлению аптичей, нижние челюсти Uraloceras, несомненно, следует классифицировать как анаптихи.
В то же время преждевременно считать дискуссию о существовании аптичей в палеозое закрытой. Многие сообщения о находках аптичей в палеозое, как и в случае с упомянутым выше пермским образцом, явно ошибочны (например, образцы, описанные Куесом (1983), на самом деле являются двустворчатыми моллюсками). Однако в 1841 году пара аптичей была описана из известняка Эйфель (эйфельский/живетский, средний девон, d'Archiac & de Verneuil, 1841: табл. XXVI. рис. 9). Хольцапфель (1899) усомнился в возрасте этой находки, но подтвердил, что он находил аналогичные образцы из сланцев Бюдесхайма (франский, верхний девон). Эти аптихи удивительно напоминают франских анаптихов формального рода Cardiocaris, которые принадлежали к гониатитидным аммоноидам (см. Мироненко 2021). Другая пара aptichi была описана Харпером (1989) из каменноугольного известняка Эймс в западной Пенсильвании (см. Harper 1989, fig. 2). Единичные находки всегда оставляют место для сомнений, однако среди "проточелюстей" предков аммоноидов - ортоцерид (формальные роды Aptychopsis, Discinocaris и Peltocaris) существовали как одно-, так и двухстворчатые формы (Мироненко 2021). Следовательно, нельзя исключать, что некоторые эволюционные линии с двустворчатыми нижними челюстями могли существовать среди ранних аммоноидов, но эта тема требует дальнейших исследований.
Экологические последствия
Судя по строению челюстного аппарата (см. рис. 6), уралоцерасы были либо активными хищниками, либо падальщиками. Об этом свидетельствуют нижние челюсти с толстым передним краем, хорошо сохранившиеся в ископаемом состоянии, и заостренные верхние челюсти. Хотя у пермских аммоноидов в челюстях не было кальцитовых элементов (они появились только в ранней юре, см. Мироненко и Гуляев 2018), это не могло помешать им быть активными хищниками, поскольку современные кальмары и осьминоги также не имеют таких минерализованных элементов в клювах, но способны охотиться даже на таких животных. хорошо защищенная добыча, такая как крабы и другие ракообразные. Трудно сказать, на каких животных охотились уралоцерасы, однако, исходя из редкости донной фауны в Дивьинской формации, наиболее вероятно, что они ловили свою добычу в толще воды. Скорее всего, их добычей были рыбы, а также головоногие моллюски небольшого размера, такие как другие аммоноиды и псевдортоцериды.
Многие из нижних челюстей Uraloceras, изученных здесь, имеют некоторые повреждения (рис. 2A,C, D), но почти все они, скорее всего, были вызваны неравномерным уплотнением осадка и посмертными деформациями. Однако в одном случае (образец K-R/13 – рис. 3B), в задней части челюсти слева от центра имеется широкое полукруглое отверстие, образование которого не может быть объяснено посмертными деформациями. Поскольку фрагмент челюсти, соответствующий этому отверстию, отсутствует, а отложения, в которых был погребен образец, образовались в условиях низкой энергии без каких-либо сильных течений, это отверстие, скорее всего, является результатом нападения хищника или действий падальщиков. За исключением двух образцов (K-R/1, см. рис. 5А, и плохо сохранившегося K-R/24), все нижние челюсти, исследованные здесь, были найдены отделенными от раковин аммоноидов и от соответствующих верхних челюстей. Это говорит о том, что челюсти уже были отделены от панцирей во время плавания в толще воды (см. Klug et al., 2021) либо в результате разложения мягких тканей, либо в результате деятельности хищников. Вышеупомянутое повреждение на одной из нижних челюстей подтверждает, что аммоноиды Uraloceras иногда могли становиться добычей хищников, включая особей примерно того же размера, что и сами аммоноиды.
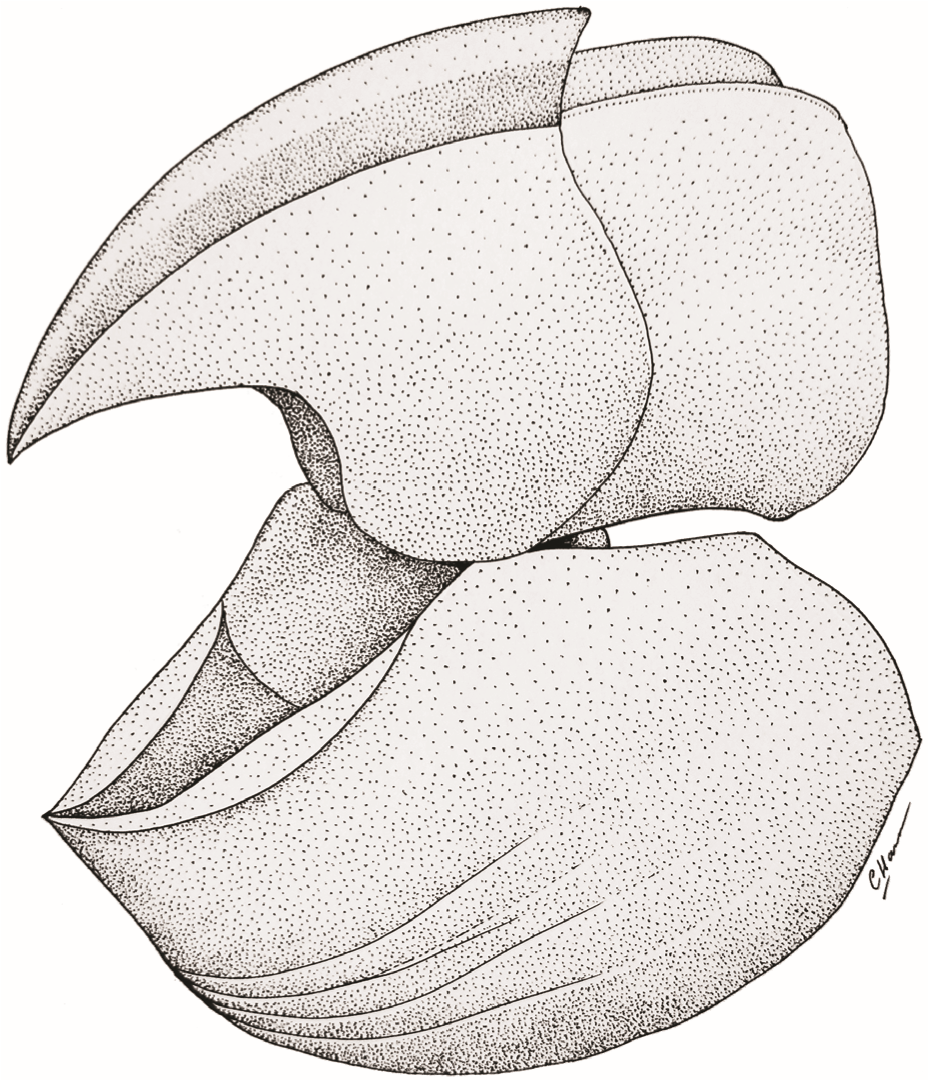
Рис. 6. Общая форма челюстного аппарата Uraloceras. Обе челюсти показаны здесь одинакового размера, хотя реальное соотношение их размеров может отличаться. Выводы
Челюсти головоногих моллюсков из дивьинской свиты артинского этапа (приуралье, Нижняя пермь), описанные здесь, скорее всего, принадлежали гониатитидным аммоноидам рода Uraloceras. Нижние челюсти типичные анаптичные, широкие и выпуклые, с гладкой или слегка ребристой внешней пластинкой и относительно большой внутренней пластинкой. В центральной части внутренней ламели расположена трапециевидная плоская область. Пластинки срослись друг с другом спереди, образуя заостренный кончик челюсти и утолщенные ‘плечики’ по ее краям. По середине наружной пластинки одного из образцов проходит борозда, чем-то напоминающая симфиз аптихов позднемезозойских аммоноидов. Верхние челюсти Uraloceras меньше соответствующих нижних челюстей, они узкие и имеют заостренные кончики. Как нижняя, так и верхняя челюсти были полностью органическими, лишенными известковых элементов. Из-за того, что верхняя челюсть имеет большой заостренный капюшон, нижние челюсти Uraloceras следует классифицировать как так называемый нормальный тип челюстей аммоноидов (Tanabe et al., 2015). Однако отсутствие скульптуры, состоящей из частых тонких ребер и линий роста, которые характерны для челюстей более древних каменноугольных гониатитов (Догужаева и др., 1997, рис. 1; Догужаева 1999: таблица 1, рис. 1,3), сближает челюсти Uraloceras со строением челюстного аппарата триасовых аммоноидов. Наличие центральной бороздки у одного экземпляра и тот факт, что эти нижние челюсти иногда складываются вдоль центральной линии, позволяют нам предположить, что какой-то гибкий симфиз мог появиться в челюстях аммоноидов уже в пермском периоде. Позже, в ранней юре, это привело к появлению челюстей типа aptychus.
Судя по заостренной форме обеих челюстей, представители рода Uraloceras, скорее всего, были активными хищниками. Тем не менее, поскольку в их челюстях отсутствовали укрепляющие известковые элементы, они охотились на добычу, у которой не было прочной внешней оболочки. Как свидетельствует находка поврежденной нижней челюсти, сами уралоцерасы также могли стать жертвами хищников, сравнимых с ними по размерам.
Признание. – Мы очень благодарны анонимным рецензентам и ответственному редактору Питеру Дойлу за их полезные и плодотворные комментарии к рукописи. Мы благодарим Михаила Рогова (ГИН РАН, Москва, Россия) за полезные и ценные комментарии к черновой версии рукописи. References Archiac, E.J.A.D’ & De Verneuil, E.P. 1842: On the fossils of the older deposits in the Rhenish provinces, preceded by a general survey of the fauna of Palaeozoic rocks, and followed by a tabular list of the organic remains of the Devonian System in Europe. Transactions of the Geological Society of London 6, 303–410. Bandel, K. 1988: Operculum and buccal mass of ammonites. In Wiedmann, J. & Kullmann, J. (eds): Cephalopods—Present and Past, 653–678. Schweizerbart, Stuttgart. Clarke, J.M. 1882: New phyllopod crustaceans from the Devonian of Western New York. American Journal of Science 23, 477–478. Closs, D. 1960: Contribuição ao estudo dos Aptychi (CephalopodaAmmonoidea) do Jurássico. Escola de Geologia de Pórto Alegre, Publicaçao Especial 2, 1–67. Closs, D. 1967: Gonatiten mit Radula und Kieferapparat in der Itararé-Formation von Uruguay. Paläontologische Zeitschrift 41, 19–37. Doguzhaeva, L.A. 1999: Beaks of the Late Carboniferous ammonoids from the Southern Urals. In: Rozanov, A.Y. & Shevyrev, A.A. (eds): Fossil Cephalopods: Recent Advances in Their Study, 68–87. Russian Academy of Science, Paleontological Institute, Moscow [in Russian]. Doguzhaeva, L.A., Mapes, R.H. & Mutvei, H. 1997: Beaks and radulae of Early Carboniferous goniatites. Lethaia 30, 305–313. Frederiks, G.N., 1915. Late Paleozoic faunas from the vicinity of Krasnoufimsk. Transactions of the Geological Committee 109, 1–115. Frye, C.J. & Feldmann, R.M. 1991: North American Late Devonian cephalopod aptychi. Kirtlandia 46, 49–71. Harper, J.A. 1989: Occurrence of a cephalopod aptychus in the Ames Limestone (Virgilian) of western Pennsylvania. Journal of Paleontology 63, 119–121. Holzapfel, E. 1899: Die Cephalopoden des Domanik im sedlichen Timan. Memoires du Comite Geologique 12, 1–56. Jones, P.J. & Olempska, E. 2013: The ostracode suborder Eridostracina Adamczak, 1961. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 267, 341–352. Karpinsky, A.P. 1890: On Ammonea of the Artinskian stratum and on some Carboniferous forms similar to them. Zapiski St. Petersburg Mineralicheskogo Obshchestva 2, 32–151. [in Russian] Karpinsky, A.P. 1891: Zur Ammoneen-Fauna der ArtinskStufe. Bulletin de l’Académie Impériale des Sciences de SaintPétersbourg, Nouvelle Serie II 2, 139–154. Keupp, H., Hoffmann, R., Stevens, K., & Albersdörfer, R. 2016: Key innovations in Mesozoic ammonoids: the multicuspidate radula and the calcified aptychus. Palaeontology 59, 775–791. Klug, C., Davesne, D., Fuchs, D., & Argyriou, T. 2020: First record of non-mineralized cephalopod jaws and arm hooks from the latest Cretaceous of Eurytania, Greece. Swiss journal of Palaeontology, 139, 1–13. Klug, C., Frey, L., Korn, D., Jattiot, R., & Rücklin, M. 2016: The oldest Gondwanan cephalopod mandibles (Hangenberg Black Shale, Late Devonian) and the mid Palaeozoic rise of jaws. Palaeontology, 59, 611–629. Klug, C., Kröger, B., Vinther, J., Fuchs, D. & De Baets, K. 2015: Ancestry, origin and early evolution of ammonoids. In Klug, C., Korn, D., De Baets, K., Kruta, I. & Mapes, R.H. (eds): Ammonoid Paleobiology: From Macroevolution to Paleogeography, 3–24. Springer, Dordrecht. Klug, C., Landman, N.H., Fuchs, D., Mapes, R.H., Pohle, A., Guériau, P., Reguer, S. & Hoffmann, R. 2019: Anatomy and evolution of the first Coleoidea in the Carboniferous. Communications Biology 2, 1–12. Klug, C., Schweigert, G., Tischlinger, H., & Pochmann, H. 2021: Failed prey or peculiar necrolysis? Isolated ammonite soft body from the Late Jurassic of Eichstätt (Germany) with complete digestive tract and male reproductive organs. Swiss Journal of Palaeontology 140, 1–14. Korn, D. 2004: Late Famennian cephalopods from the Refrath 1 Borehole (Bergisch Gladbach-Paffrath Syncline; ArdennesRhenish Massif Germany). Courier Forschung-Institut Senckenberg, 251, 175–183. Krotov, P.I. 1885: Artinskian Stage. Geological-paleontological monograph on the Artinskian sandstone. Proceedings of the Society of Naturalists of the Kazan University 13, 1–314 [In Russian]. Kruta, I., Landman, N.H., Mapes, R. & Pradel, A. 2014: New insights into the buccal apparatus of the Goniatitina; palaeobiological and phylogenetic implications. Lethaia 47, 38–48. Kues, B.S. 1983: Cephalopod aptychi from Los Moyos Limestone, Madera Group (Middle Pennsylvanian),near Albuquerque, New Mexico. New Mexico Geology 5, 78–80. Landman, N.H., & Davis, R.A. 1988: Jaw and crop preserved in an orthoconic nautiloid cephalopod from the Bear Gulch Limestone (Mississippian, Montana). New Mexico Bureau of Mines and Mineral Resources, Memoir 44, 103–107. Landman, N.H., Garb, M.P., Rovelli, R., Ebel, D.S., & Edwards, L.E. 2012: Short-term survival of ammonites in New Jersey after the end-Cretaceous bolide impact. Acta Palaeontologica Polonica, 57, 703–715. Landman, N.H., Mapes, R.H., & Cruz, C. 2010: Jaws and soft tissues in ammonoids from the Lower Carboniferous (Upper Mississippian) Bear Gulch Beds, Montana, USA. In: Tanabe, K., Sasaki, T., Shigeta, Y., Hirano, H. (eds): Cephalopods — Present and Past, 147–153. Tokai University Press, Tokyo. Lehmann, U. 1967: Ammoniten mit Kieferapparat und Radula aus Lias-Geschieben. Paläontologische Zeitschrift 41, 38–45. Manda, Š., & Turek, V. 2011: Late Emsian Rutoceratoidea (Nautiloidea) from the Prague Basin, Czech Republic: morphology, diversity and palaeoecology. Palaeontology 54, 999–1024. Mapes, R.H. 1987: Upper Paleozoic cephalopod mandibles: frequency of occurrence, modes of preservation, and paleoecological implications. Journal of Paleontology 61, 521–538. Mapes, R.H., Weller, E.A., & Doguzhaeva, L.A. 2010: Early Carboniferous (Late Namurian) coleoid cephalopods showing a tentacle with arm hooks and an ink sac from Montana, USA. In: Tanabe, K., Shigeta, Y., Sasaki, T. & Hirano, H. (eds): Cephalopods – Present and Past, 155–170. Tokai University Press, Tokyo. Meek, F.B., Hayden, F.V. 1864: Paleontology of the Upper Missouri. Smithonian Contributions to Knowledge 172, 1–135. Mikulash, R. & Dronov, A.V. 2006: Paleoichnology—Introduction to the Study of Trace Fossils, Geologicheskiy Institut Cheshskoi Akademiia Nauk, Praga. 122 p. Mironenko, A.A. 2021: Early Palaeozoic Discinocarina: a key to the appearance of cephalopod jaws. Lethaia 54, 457–476. Mironenko, A., & Gulyaev, D. 2018: Middle Jurassic ammonoid jaws (anaptychi and rhynchaptychi) from Dagestan, North Caucasus, Russia. Palaeogeography, Palaeoclimatology, Palaeoecology 489, 117–128. Naugolnykh, S.V. 2016: Paleontologiya Krasnoufimska (Paleontology of Krasnoufimsk), Moscow: Media-Grand, 72 p. Naugolnykh, S.V. 2018: Artinskian (Early Permian) Sea Basin and Its Biota (Krasnoufimsk, Cis-Urals). Stratigraphy and Geological Correlation 26, 734–754. Ruzhentsev, V.E. & Shimanskiy, V.N. 1954: Nizhnepermskie svernutye i sognutye Nautiloidei yuzhnogo Urala. Akademiya Nauk SSSR, Trudy Paleontologicheskogo Instituta 50, 152 p. Spinosa, C. 1968: The Xenodiscidae, Permian Otoceratacean Ammonoids. Unpublished Ph.D. dissertation, University of Iowa. Tanabe, K., Kruta, I. & Landman, N.H. 2015: Ammonoid buccal mass and jaw apparatus. In Klug, C., Korn, D., De Baets, K., Kruta, I. & Mapes, R.H. (eds): Ammonoid Paleobiology: From Anatomy to Ecology. Topics in Geobiology 43, 439–494. Springer, Dordrecht Tanabe, K. & Mapes, R.H. 1995: Jaws and radula of the Carboniferous ammonoid Cravenoceras. Journal of Paleontology 69, 703–707. Trauth, F. 1927: Aptychenstudien, I. Annalen des Naturhistorischen Museums in Wien 41, 171–259. Woodward, E. 1882: Note on Ellipsocaris Dewalquei, a new Phyllopod Crustacean Shield from the Upper Devonian of Belgium. Geological Magazine 9, 444–446. Woodward, H. 1885: On some Palæozoic Phyllopod shields, and on Nebalia and its allies. Geological Magazine 2, 345–352. Zakharov, Yu. D. 1979: New finds of rhyncholites, anaptychi, aptychi and remains of cephalopod radulae at USSR territory. In: M.N. Gramm (ed): Fossil Invertebrates of the Far East, 80–99. Vladivostok [in Russian].
|
|||||||||
|
|
Последнее изменение этой страницы: 2024-06-27; просмотров: 13; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.95.167 (0.009 с.) |