Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
повышенная соленость в придонном слое воды.Содержание книги
Поиск на нашем сайте
Нижняя и верхняя челюсти раннепермских гониатидных аммоноидов А.А. Мироненко и С.В. Наугольных Аммоноиды — головоногие моллюски с внешними раковинами, существовавшие от раннего девона до конца мелового периода, — имели хорошо развитые челюсти. В ходе эволюции аммоноидов возникло несколько различных типов их челюстного аппарата, изучение которых представляет несомненный интерес, поскольку позволяет исследователям делать выводы о стратегиях питания аммоноидов и их положении в трофических цепочках. Тем не менее, нет результатов, касающихся эволюции аммоноидных челюстей во время перми. Здесь мы опишем коллекцию из почти тридцати челюстей головоногих моллюсков из Дивжинской свиты (Артинский этап, Цисуральский, Нижнепермский), из Свердловской области России. Скорее всего, эти челюсти принадлежат гониатидному аммоноиду Uraloceras, наиболее распространенному головоногому моллюску в дивжинской (дивьской) формации. Нижние челюсти Uraloceras представляют собой типичные аммоноидные анаптыхи, которые имеют округлую, широкую и выпуклую форму с гладкой или слегка ребристой поверхностью. Они имеют большую внутреннюю пластинку с трапециевидной платформой в центральной части. Одна из челюстей несет возможный след укуса хищника или падальщика. Верхние челюсти, описанные здесь впервые, немного меньше нижних, их форма узкая и заостренная. Первоначально обе челюсти были полностью органичными без известковых элементов. Отсутствие скульптуры, состоящей из частых ребер и линий роста, характерных для более древних каменноугольных гониатидных челюстей, делает челюсти уралоцер ближе к строению челюстного аппарата триасовых аммоноидов. Судя по заостренной форме кончиков обеих челюстей, уралоцеры были активными хищниками. □ Permian, Cephalopoda, Ammonoidea, jaw apparatus, anaptychi, Artinskian Aleksandr A. Mironenko ✉ [paleometro@gmail.com] and Serge V. Naugolnykh [naugolnykh@list.ru], Geological Institute of Russian Academy of Sciences, Pyzhevski Lane 7, 119017 Moscow, Russia; manuscript received on 10/11/2021; manuscript accepted on 01/05/2022, manuscript published on 19/12/2022 in Lethaia 55(4). Аммоноиды, как и современные головоногие моллюски, имели хорошо развитый челюстной аппарат, который состоял из пары челюстей - верхней и нижней, и радулы, заключенной между ними. Каждая челюсть состояла из двух пластинок – внутренней и внешней. Соотношение размеров этих пластинок у головоногих моллюсков различно, но у аммоноидов наружная пластинка всегда больше внутренней в нижней челюсти, тогда как в верхней челюсти, наоборот, внутренняя пластинка обычно больше (см. Tanabe et al., 2015). Находки элементов челюстного аппарата аммоноидов известны из отложений всех периодов, когда существовали аммоноиды: от девона до конца мелового периода или даже до самого начала палеогена (Landman et al., 2012; Tanabe et al., 2015). Изучению челюстного аппарата аммоноидов посвящено много публикаций, однако палеозойская стадия его эволюции до сих пор изучена меньше, чем мезозойская (см. обзор Tanabe et al., 2015 и Keupp et al., 2016). Из пяти типов челюстных аппаратов аммоноидов, идентифицированных на данный момент, четыре относятся к мезозойским аммоноидам и только один - к палеозойским (Tanabe et al., 2015).
Аммоноиды возникли в конце раннего девона, в эмсе (Klug et al., 2015). Что касается палеозоя, то находки их челюстей известны из девонских, каменноугольных и пермских отложений. Однако количество этих находок в разные периоды и уровень их осведомленности весьма противоречивы. Широко признано, что древнейшие известные на сегодняшний день челюсти аммоноидов происходят из позднедевонских (франских) отложений (Tanabe et al., 2015; Klug et al., 2016). Действительно, наибольшее количество челюстей девонских аммоноидов происходит из франского периода и принадлежит к семейству Gephuroceratidae (Woodward 1885; Trauth 1927). Меньшее количество челюстей аммоноидов описано из фаменских отложений (Fry & Feldmann 1991; Korn 2004; Klug et al. 2016). Первоначально нижние челюсти девонских аммоноидов рассматривались как филлоподы-ракообразные и были включены в подотряд Discinocarina (например, Woodward 1882). Один из родов дискинокаринов, Lisgocaris, был описан Кларком (1882) из отложений, которые, по современным данным, относятся к живетской стадии среднего девона (см. Frye & Feldmann 1991; Jones & Olempska 2013). Таким образом, получается, что самые древние челюсти аммоноидов известны из живетии. Нет никаких сомнений в том, что эмсийские и эйфелевские челюсти аммоноидов также должны были существовать (Klug et al., 2016), и отсутствие их находок, скорее всего, связано с тафономическими причинами (см. Мироненко 2021). Находки челюстей аммоноидов довольно многочисленны в отложениях каменноугольного периода (как миссисипских, так и пенсильванских) (Клосс 1960, 1967; Мэйпс 1987; Бандель 1988; Танабе и Мэйпс 1995; Догужаева и др. 1997; Догужаева 1999; Ландман и др. 2010; Крута и др. 2014). Находки челюстей аммоноидов триасового периода также очень многочисленны, что отражено в большом количестве публикаций, посвященных этим окаменелостям (см. обзор Tanabe et al., 2015). Однако знания об аммоноидных челюстях пермского периода очень скудны. Челюсти пермских аммоноидов были описаны на сегодняшний день всего несколько раз. Впервые они были описаны в конце 19 века на артинском ярусе Пермской области России (Кротов, 1885). Одна челюсть аммоноида, странно идентифицированная как aptychus, была описана из артинских отложений в окрестностях города Красноуфимска А.П. Карпинским (1890, 1891). Фредерикс (1915) также изобразил образец из нижней перми в Красноуфимске и интерпретировал его как aptychus, но мы повторно изучили эту запись и пришли к выводу, что он не имеет отношения к челюстям головоногих моллюсков. Одна нижняя челюсть аммоноида (anaptychus) была описана из камеры тела аммоноида Paraceltites rectangularis Miller из пермских (капитанских) отложений Мексики в неопубликованной докторской диссертации Спинозы (1968). Несколько мелких пермских нижних челюстей (anaptychi) были также описаны Ю.Д. Захаровым (1979) как новый официальный вид Anaptychus ruzhencevi. Однако эти данные очень ограничены, количество опубликованных на данный момент образцов очень незначительно, и на самом деле мы можем только представить, как изменялись челюсти аммоноидов на протяжении пермского периода, сравнивая более старые образцы каменноугольного периода и более молодого триаса друг с другом. Сами челюсти пермских аммоноидов до сих пор оставались практически неисследованными. Верхние челюсти пермских аммоноидов никогда не были описаны.
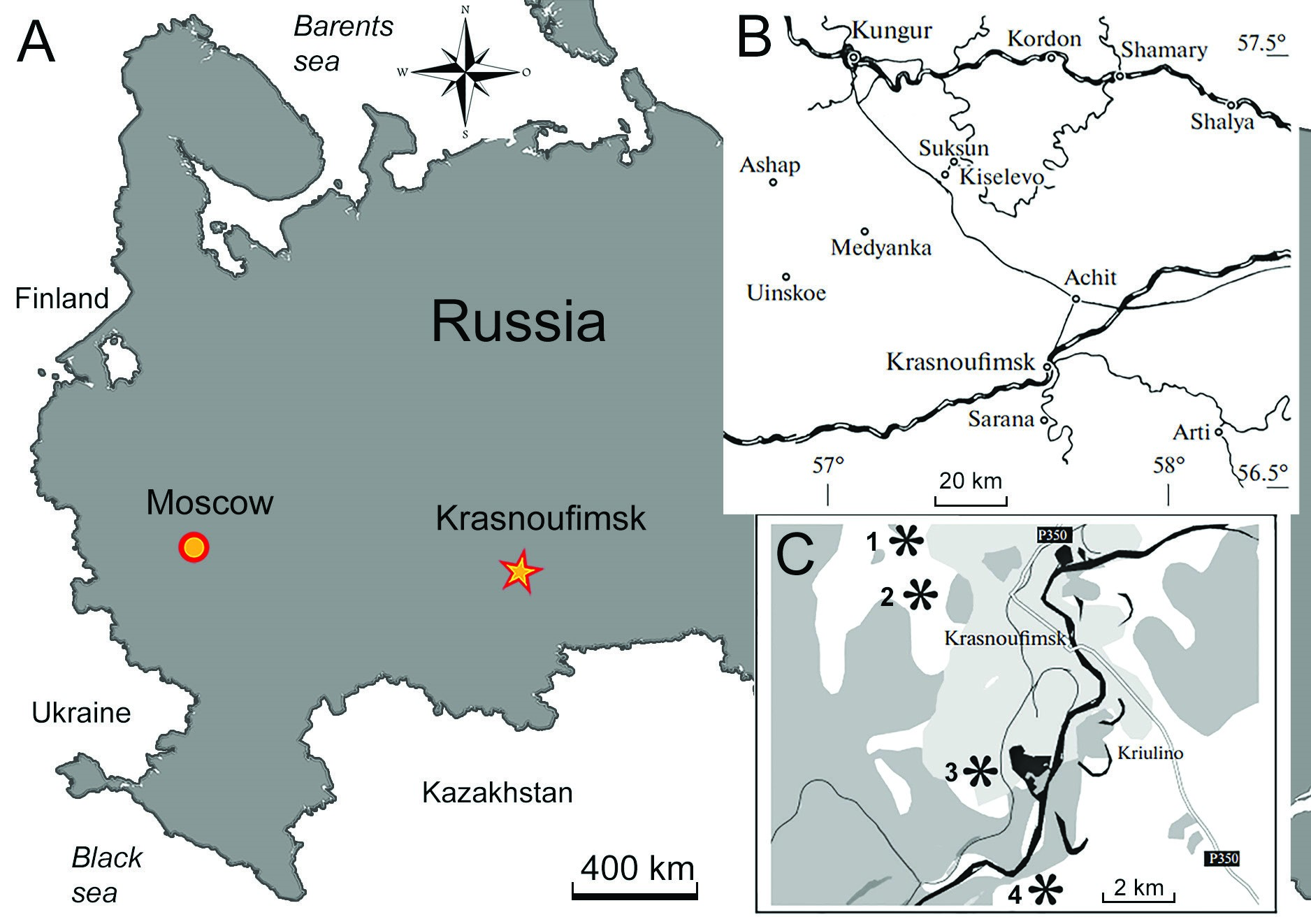
Рис. 1. Карты населенных пунктов. А, Красноуфимск на карте России. Б, окрестности Красноуфимска. C, населенные пункты, где были найдены челюсти аммоноидов: 1, Красноуфимские ключи; 2, За Селекцией; 3, Соболя; 4, Черкасова. В этой статье мы описываем коллекцию челюстей пермских аммоноидов, состоящую из 28 экземпляров. Все евреи происходят из дивьинской формации артинского яруса (приуралье, Нижняя пермь), обнажившейся в окрестностях города Красноуфимск Свердловской области, Россия (рис. 1). Все исследованные образцы находятся в коллекции Государственного геологического музея имени В.И. Вернадского (Москва). Коллекция включает в себя как хорошо сохранившиеся нижнюю, так и верхнюю челюсти. Это самый большой набор челюстей пермских аммоноидов, изученных на сегодняшний день. Отличная сохранность многих экземпляров позволила нам изучить первоначальную форму челюстей и сделать некоторые предположения об экологии их возможных хозяев — гониатитидных аммоноидов рода Uraloceras Ruzhencev, 1936. Геологические условия Челюсти головоногих моллюсков были собраны одним из нас (SVN) в четырех населенных пунктах в окрестностях города Красноуфимск Свердловской области, Россия (рис. 1). Это Красноуфимские Ключи, откуда взято большинство экземпляров (22 челюсти), Соболя (3), За Селекцией (2) и Черкасово (1) (см. Наугольных 2018). Все челюсти были найдены в голубовато-серых пластинчатых мергелях дивьинской свиты, саргинском горизонте артинского яруса (приуралье, пермь). Помимо челюстей головоногих моллюсков, дивьинская формация (Наугольных 2016, 2018; также известная как Дивьяская формация, напр. Лебедев 2009) в районе города Красноуфимска содержит панцири трилобитов, многочисленные останки рыб (в том числе зубчатые завитки Helicoprion bessonowi Karpinsky), конуляриид и крупные раковины наутилоидов и аммоноидов. Следует отметить, что арагонитовые слои раковин головоногих моллюсков Дивьинской свиты полностью растворены, тогда как их внутренние формы хорошо сохранились. В то же время фосфатные окаменелости и обуглившееся органическое вещество растительных остатков и челюстей головоногих моллюсков хорошо сохранились в этих слоях. Из-за находок зубчатых мутовок Helicoprion Karpinsky дивьинскую формацию иногда называют "мергелевой формацией Helicoprion" (Наугольных, 2016). Однако наиболее распространенным типом окаменелостей в Дивьинской формации являются окаменелые следы – извилистые следы ползающих червеобразных донных животных, которые жили в донном иле или на его поверхности (например, Наугольных 2016, рис. 2). Эти следы окаменелостей можно отнести к формальному роду Helminthoides (Микулаш и Дронов, 2006).
Артинский бассейн в районе Красноуфимска был частью Приуральского бассейна Артинского моря, который был соединен с Бореальным океаном на севере и морем Тетис на юге (Наугольных 2018). Редкость находок донных животных с преобладанием ихнофоссилий и остатков пелагических организмов (головоногих моллюсков и рыб) указывает на то, что эти отложения формировались на относительно значительной глубине (Наугольных 2016, 2018). Кроме того, нельзя исключать наличие неблагоприятных факторов, таких как дефицит кислорода или
Рис. 2. Нижние челюсти, на которых обнажена только одна наружная пластинка или ее отпечаток. A, образец KR/9. B, K-R/8. C, K-R/5. D, K-R/11. E, K-R/21. F, K-R/12. Линейки шкалы: 5 мм. Методы и материалы
Изученные здесь образцы были исследованы как с помощью оптического бинокулярного микроскопа, так и под сканирующим электронным микроскопом (SEM; TESCAN//VEGA с выдвижным детектором BSE) в Палеонтологическом институте Российской академии наук (PIN РАН). Однако исследование на СЭМ не принесло существенных результатов из-за недостаточной сохранности органических слоев челюстей. Среди 28 образцов челюстей, изученных здесь, 23 идентифицированы как нижние челюсти (два из них – предварительно) и пять идентифицированы как верхние челюсти (один из них - предварительно). Большинство находок представляют собой слепки или отпечатки одной или двух пластинок челюстей, образованных мелкозернистым осадком. Органические слои челюстей обычно отсутствуют. Однако в некоторых образцах нижних челюстей обуглившееся органическое вещество сохраняется в утолщенных частях вдоль переднего края, по бокам кончика, на стыке наружной и внутренней пластинок, а в некоторых случаях даже в виде тонкого слоя на поверхности оттисков внешних пластинок. Нижние челюсти можно разделить на три группы в зависимости от степени сохранности.
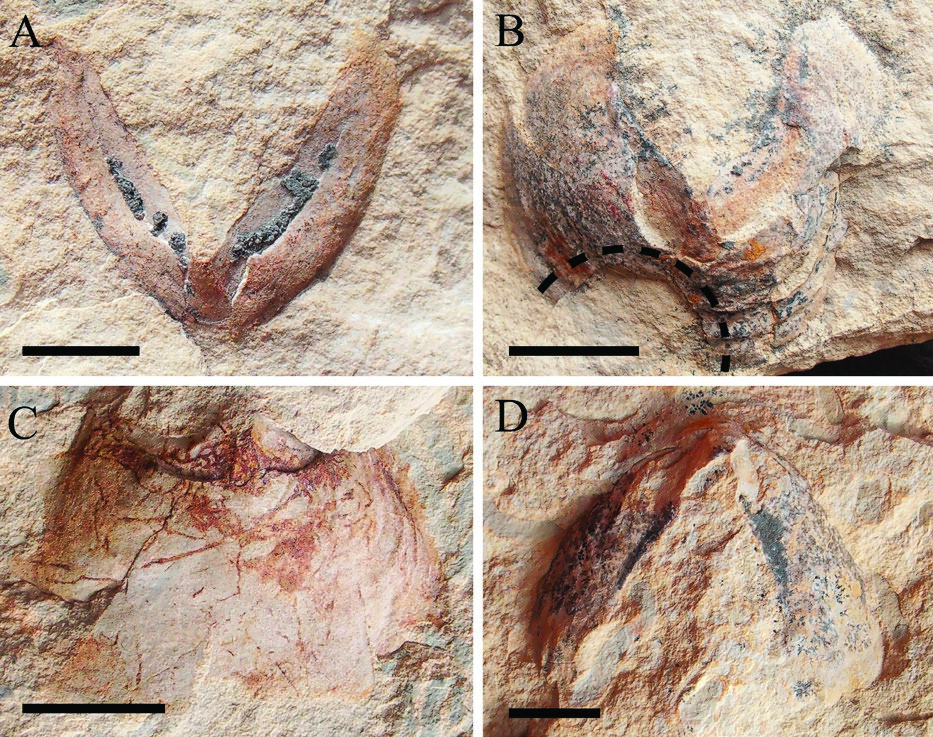
Первая и наиболее многочисленная группа включает образцы, у которых сохранилась или видна только одна наружная пластинка или ее отпечаток (рис. 2). Такие образцы позволяют нам понять общую форму этих нижних челюстей: они были широкими, с закругленным задним краем и заостренным передним кончиком. Первоначально челюсти были выпуклыми, некоторые из них были сплющены во время диагенетического уплотнения осадка и растрескались по заднему краю с многочисленными трещинами (например, образец K-R/9, см. рис. 2A). Такая форма типична для палеозойских и мезозойских аммоноидных анаптичей (см. Tanabe et al., 2015). В подавляющем большинстве челюстей внешняя пластина выглядит монолитной, но в образце K-R/11 (рис. 2D) вдоль его центральной части проходит тонкая канавка. Эта борозда чем-то напоминает симфиз аптихов мезозойских аммонитов. В некоторых случаях наружная пластинка нижних челюстей имеет слегка волнистую форму с широкими пологими ребрами (рис. 2Б). Тем не менее, трудно сказать, является ли это его первоначальной формой или результатом посмертных деформаций. Передние кончики челюстей острые. Вторая группа включает образцы, на которых также виден отпечаток внутренней пластинки нижней челюсти вместе с внешней пластинкой (рис. 3). Внутренняя пластинка широкая и длинная, по крайней мере, вдвое меньше длины внешней пластинки. В центральной части внутренней пластинки имеется уплощенная трапециевидная платформа, которая сужается к кончику челюсти и расширяется кзади (рис. 3А, Б, Г). Рис. 3. Нижние челюсти, на которых также виден отпечаток внутренней пластинки нижней челюсти вместе с внешней пластинкой. A, образец K-R/7. B, K-R/13. C, K-R/2. D, K-R/6. Линейки шкалы: A, D, 5 мм; B, C. 1 см. Рис. 4. Верхние челюсти. A, образец KR/22. B, K-R/26. C, K-R/19. D, K-R/27. Линейки шкалы: 5 мм. Третья группа изученных образцов включает фрагментарно сохранившиеся челюсти, интерпретация которых часто затруднена из-за неполной сохранности. Мы не приводим их изображений, поскольку они заслуживают упоминания только в смысле подсчета количества находок, но не представляют интереса для реконструкции строения челюстей. Верхние челюсти (рис. 4) намного уже и острее нижних. В большинстве случаев видна только их наружная пластинка с удлиненным передним концом и боковыми выступами (рис. 4С,D). Размер образцов верхней челюсти, в среднем, меньше размера нижних, и хотя не было найдено ни одной пары челюстей (т.е. как верхней, так и нижней челюстей одного и того же животного), можно предположить, что верхние челюсти были несколько меньше по размеру, чем нижние. В то же время разделение внутренних пластинок на две удлиненные половины, характерное для верхних челюстей мезозойских аммонитов, не было обнаружено в изученных здесь образцах. Как верхняя, так и нижняя челюсти пермских головоногих моллюсков, изученные здесь, не содержат признаков известковых элементов. В то время как кальцит и арагонит из раковин моллюсков не сохранились в Дивьинской формации, внутренние формы и отпечатки раковин сохранились с превосходной детализацией. Поскольку исследованные челюсти не содержали никаких отпечатков известковых ринхолитов или конхоринхов, они, скорее всего, были полностью органическими.
|
|||||||||
|
|
Последнее изменение этой страницы: 2024-06-27; просмотров: 5; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.21.44.115 (0.009 с.) |