Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Эволюция полового размножения у дрожжей: надстройка меняется, базис остаетсяСодержание книги
Поиск на нашем сайте
Эволюция полового размножения у дрожжей: надстройка меняется, базис остается
25.02.14 | Генетика, Биология развития, Эволюция, Александр Марков | Комментировать
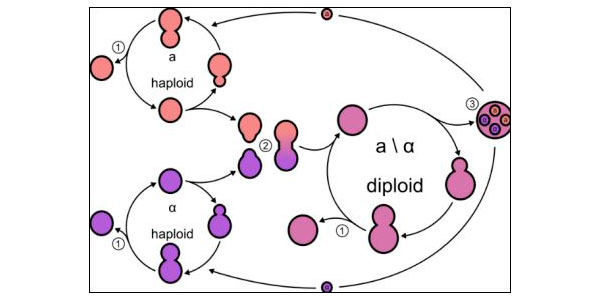
Рис. 1. Схема жизненного цикла пекарских дрожжей Saccharomyces cerevisae. 1 — почкование, 2 — конъюгация (спаривание), 3 — образование четырех гаплоидных спор из диплоидной клетки в результате мейоза. Схема с сайта www.wynboer.co.za Американские биологи обнаружили, что генные сети, регулирующие половое размножение, по-разному устроены у разных видов дрожжей. У обычных пекарских дрожжей и ряда других видов спаривание (слияние двух гаплоидных клеток в одну диплоидную) и мейоз (образование гаплоидных спор из диплоидной клетки) разнесены во времени, а в жизненном цикле преобладает диплоидная фаза. У других видов мейоз происходит вскоре после спаривания, и поэтому в жизненном цикле преобладает гаплоидная фаза. Как выяснилось, в первом случае спаривание и мейоз управляются двумя независимыми генетическими программами, а во втором эти программы слиты воедино, причем белки-регуляторы, необходимые для спаривания, стали необходимыми также и для мейоза, и наоборот. По-видимому, слияние управляющих программ происходило независимо в разных группах дрожжей, когда им становилось выгодно проводить большую часть жизни в гаплоидном состоянии. У организмов, размножающихся половым путем, в жизненном цикле обязательно присутствуют два особых события – оплодотворение, в результате которого два гаплоидных генома объединяются в одной клетке, и редукционное деление (мейоз), в ходе которого из диплоидной клетки получаются гаплоидные. Иногда эти события тесно связаны и следуют одно за другим, а иногда — разнесены во времени и происходят более или менее независимо. Первый вариант характерен, например, для животных (у которых оплодотворение следует за мейозом), а также для многих одноклеточных, проводящих большую часть жизни в гаплоидном состоянии (в этом случае, наоборот, мейоз происходит сразу после оплодотворения). Пример второго варианта — пекарские дрожжи Saccharomyces cerevisae, классический лабораторный объект. Диплоидные клетки S. cerevisae могут долго размножаться почкованием, производя дочерние диплоидные клетки. В определенных условиях (например, при голодании) диплоидная клетка претерпевает мейоз и производит четыре гаплоидные споры. Из спор вырастают гаплоидные дрожжевые клетки, которые тоже могут размножаться почкованием, однако они, в отличие от диплоидных клеток, способны еще и к спариванию (конъюгации). Гаплоидные дрожжи делятся на два пола (a и α), причем спариваться могут только разнополые клетки. В результате спаривания получается диплоидная клетка, и цикл продолжается (рис. 1). Таким образом, уS. cerevisae между мейозом и оплодотворением могут быть долгие периоды бесполого размножения, а сами эти два события друг с другом напрямую не связаны и регулируются разными генами. По-другому устроен жизненный цикл родственника пекарских дрожжей, патогенного гриба Candida lusitaniae. У этого вида большая часть жизни проходит в гаплоидной фазе, а образовавшаяся в результате конъюгации диплоидная клетка, как правило, не почкуется (или почкуется недолго) и вскоре приступает к мейозу. Микробиологи из Брауновского университета (США) решили разобраться в причинах этого различия. Они сравнили экспрессию генов S. cerevisae и C. lusitaniae во время спаривания и мейоза. У S. cerevisae наборы генов, отвечающих за эти два процесса, четко различаются. Однако у C. lusitaniae, как выяснилось, разделение труда между двумя группами генов выражено намного слабее. Во время спаривания у C. lusitaniae активируются не только гены, которые у пекарских дрожжей отвечают за спаривание (включая гены MAPK-каскада, обеспечивающего реакцию клетки на половые феромоны противоположного пола), но и некоторые гены, которые у S. cerevisae необходимы для мейоза, но в спаривании не участвуют. К числу таких «генов мейоза», активирующихся у C. lusitaniae во время спаривания, относятся SPO11, REC8 и IME2. В диплоидных клетках C. lusitaniae, собравшихся приступить к мейозу, резко (более чем в четыре раза) возрастает экспрессия 618 генов. Самое интересное, что многие из этих генов у S. cerevisaeобслуживают исключительно процесс спаривания, а в мейозе не участвуют (у S. cerevisae при мейозе возрастает экспрессия меньшего числа генов — 480). В частности, многие гены, у пекарских дрожжей и других грибов отвечающие за реагирование на половые феромоны, у C. lusitaniae почему-то активируются не только при подготовке к спариванию, но и в ходе мейоза. С другой стороны, некоторые гены, активирующиеся у S. cerevisae при спаривании или мейозе, у C. lusitaniae в этих ситуациях не активируются. Ключевой регулятор мейоза IME1 вообще отсутствует уC. lusitaniae. Полученные факты позволяют заключить, что генные сети, регулирующие спаривание и мейоз у двух видов, существенно различаются. Чтобы выяснить, имеют ли эти различия функциональное значение, авторы приступили к изучению работы отдельных генов. Ген IME2 (inducer of meiosis 2) является ключевым регулятором мейоза у S. cerevisae. Как и следовало ожидать, удаление этого гена у C. lusitaniae лишило диплоидные клетки способности к мейозу. Но это не всё: гаплоидные клетки C. lusitaniae без гена IME2 оказались неспособны к спариванию. Между тем у S. cerevisae ген IME2 на спаривание не влияет. Половые феромоны необходимы пекарским дрожжам для регуляции спаривания, но не мейоза. Оказалось, что C. lusitaniae активно производят феромоны и в ходе мейоза, и при спаривании. Ген STE12 кодирует транскрипционный фактор, играющий ключевую роль в регуляции полового поведения у S. cerevisae и многих других грибов. Его удаление у C. lusitaniae лишило клетки не только способности к спариванию (этот результат был ожидаем), но и способности к мейозу, при том что у S. cerevisae этот ген в мейозе не задействован. Вся совокупность данных говорит о том, что у S. cerevisae есть две более или менее самостоятельные генетические программы, одна из которых регулирует половое поведение (спаривание), а другая — мейоз. У родственного вида C. lusitaniae эти две программы фактически слились в одну. Авторы проверили, влияет ли удаление гена STE12 на мейоз еще у трех близких видов дрожжей: Kluyveromyces lactis, Pichia pastoris и Yarrowia lipolytica. Во всех трех случаях никакого влияния выявлено не было (как и у S. cerevisae). Это значит, что разделенность программ спаривания и мейоза, по-видимому, является исходным (предковым) признаком данной группы грибов, а их слияние у C. lusitaniae — признак новоприобретенный (рис. 2). Возможно, ключом к слиянию генетических программ послужила утрата транкрипционного фактора α2. Об этом белке известно, что он не позволяет некоторым генам, активным в гаплоидных клетках (в том числе генам MAPK-каскада, отвечающего за реагирование на половые феромоны), работать также и в диплоидной фазе жизненного цикла. Предки C. lusitaniae утратили α2, что дало возможность «генам спаривания» работать в диплоидных клетках. Это и позволило некоторым из них взять на себя новую функцию регуляторов мейоза. Слияние программ спаривания и мейоза не является уникальной особенностью C. lusitaniae. Такое же явление обнаружено у дальнего родственника рассматриваемой группы,Schizosaccharomyces pombe, чьи предки отделились от предков других дрожжей более 330 млн лет назад. У этого вида, как и у C. lusitaniae, транскрипционные факторы MAPK-каскада, необходимые для спаривания, необходимы также и для мейоза.
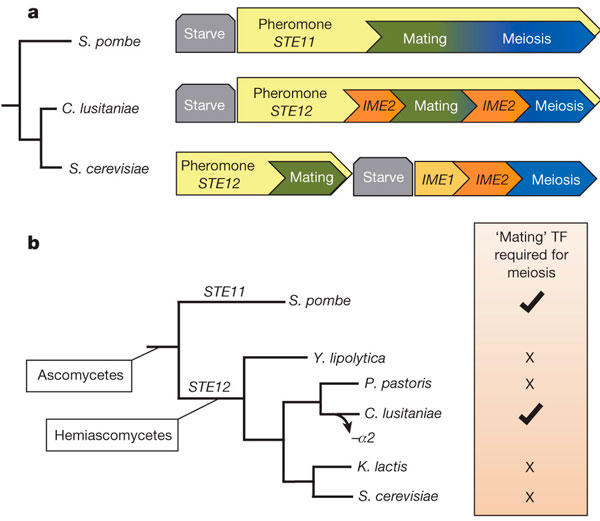
Рис. 2. Вверху (a) — схема регуляции спаривания (Mating) и мейоза (Meiosis) у некоторых видов дрожжей. У Schizosaccharomyces pombe и Candida lusitaniae оба процесса регулируются совместно, с участием половых феромонов и белков MAPK-каскада, и индуцируются голоданием. У Saccharomyces cerevisae два процесса регулируются по отдельности, а половые феромоны нужны только для спаривания. Внизу (b) — эволюционное дерево, показывающее, что совместная регуляция спаривания и мейоза возникла уS. pombe и C. lusitaniae независимо. Справа отмечены галочками виды, у которых транскрипционные факторы Ste11 и Ste12 необходимы не только для спаривания, но и для мейоза; крестиками — те, кому они нужны только для спаривания. Рисунок из обсуждаемой статьи в Nature По-видимому, слияние генетических программ спаривания и мейоза происходило в эволюции грибов неоднократно. По мнению авторов, это могло быть связано с тем, что некоторым видам почему-то становилось выгодно большую часть жизненного цикла проводить в гаплоидной фазе (хотя в чем именно состоят преимущества и недостатки гаплоидности и диплоидности — вопрос дискуссионный, см.: S. P. Otto, A. C. Gerstein, 2008. The evolution of haploidy and diploidy). Единая регуляция спаривания и мейоза у C. lusitaniae и S. pombe приводит к тому, что мейоз у них обычно происходит вскоре после образования диплоидной клетки, и поэтому большую часть жизни эти грибы проводят в гаплоидном состоянии. Напротив, у S. cerevisae и других видов с разделенными программами спаривания и мейоза в жизненном цикле преобладает диплоидная фаза. Исследование еще раз показало, что в ходе эволюции может происходить радикальная перестройка (rewiring) генных сетей, регулирующих развитие того или иного признака, хотя сам признак при этом не меняется. Другой пример — огромное разнообразие способов детерминации пола у разных организмов, хотя в итоге получается одно и то же: часть особей становится самцами, часть — самками (впрочем, к дрожжам с их «полами» а и α это как раз не относится). Источник:Racquel Kim Sherwood, Christine M. Scaduto, Sandra E. Torres & Richard J. Bennett. Convergent evolution of a fused sexual cycle promotes the haploid lifestyle // Nature. 2014. V. 506. P. 386–390. Про половое размножение дрожжей см. также: Про устройство MAPK-каскада (обеспечивающего реакцию дрожжей на половые феромоны противоположного пола) см. также:
|
||||
|
|
Последнее изменение этой страницы: 2024-06-27; просмотров: 4; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.14.254.227 (0.008 с.) |