Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Лекція 3. Основні абіотичні фактори середовища: світло, температура, вологаСодержание книги
Поиск на нашем сайте
Лекція 3. ОСНОВНІ АБІОТИЧНІ ФАКТОРИ СЕРЕДОВИЩА: СВІТЛО, ТЕМПЕРАТУРА, ВОЛОГА Світло в житті організмів. Температура в житті організмів. Волога в житті організмів. Адаптації організмів: активні, пасивні і такі, що дозволяють уникнути несприятливого впливу середовища.
Світло в житті організмів. Світло є електромагнітними хвилями різної довжини (від 0,05 до 3000 нм). Екологічне значення освітленості визначається трьома характеристиками світлового потоку: тривалістю (фотоперіодом), інтенсивністю і якістю (довжиною хвилі). Щохвилини Земля отримує 2 кал/см2 сонячної енергії (1,39 х 103дж/м2 с). Ця величина називається сонячною постійною. Спектр світла складається з областей:<150 нм – іонізуюча радіація – < 0,1%; 150-380 нм – ультрафіолетова радіація (УФ) – 1-10%;380-720 нм – видиме світло – до 50%;720-1000 нм – інфрачервона радіація (ІЧ) – до 50%. Але не вся промениста енергія сягає земної поверхні. До 19% розсіюється в атмосфері (парами і пилом, молекулами газів), близько 34% відбивається від атмосфери (від хмар) в космічний простір і лише 47% сонячної енергії сягає біосфери. Іонізуюче випромінювання майже повністю затримується верхніми шарами атмосфери. Ультрафіолетові промені (УФ) в помірних дозах стимулюють ріст і розмноження клітин, сприяють синтезу біологічно активних речовин, вітамінів, антибіотиків і тим самим підвищують стійкість до хвороб. Короткі хвилі цього випромінювання (200-320 нм) мають канцерогенну дію, але велика частина їх поглинається озоновим шаром атмосфери. До поверхні Землі доходять в основному хвилі довші за 300 нм, викликаючи у тварин синтез антирахітичного гормону D. Багато звірів вранці виносять з нор своїх дитинчат на сонці, у птахів – «сонячне купання». Передозування УФ шкідливе, особливо для поділу клітин, тому УФ використовують для дезінфекції приміщень. Як захист від зайвих доз УФ в шкірі людини і інших ссавців утворюється пігмент меланін (загар). Інфрачервоне випромінювання (ІК) сприймається організмами як тепло. Впливаючи на теплові центри нервової системи тварин, ці промені регулюють швидкість окислювальних процеси і рухові реакції відносно джерела тепла. Найбільше значення для живих організмів і функціонування всієї біосфери має видима частина спектру, що складається з прямої (27%) і розсіяної (16%) радіації. Всі промені, що впливають на рослинні організми, особливо на фотосинтез, називаються фізіологічно активною радіацією (ФАР). На світлі відбувається утворення хлорофілу і вже за його участі здійснюється фотосинтез: 6CO2 + 6H2O = С6Н12О6 + 6O2. Зеленими рослинами на суші найактивніше поглинаються червоні (650-680 нм) і синьо-фіолетові (400-500 нм) промені. У водних глибинах спектр світла змінений, взагалі відсутні червоні промені, тому у водоростей утворюються додаткові фотосинтетичні пігменти (фікобіліни), що дозволяє їм жити в морі на великій глибині. Пурпурні і зелені бактерії, які мають бактеріохлорофіли, здатні поглинати світло в довгохвильовій частині (максимуми 800—1100 нм). Це дозволяє їм існувати навіть за наявності тільки невидимих інфрачервоних: променів. На інтенсивність світла впливає кут падіння сонячних променів на земну поверхню, який змінюється залежно від широти, сезону, часу дня і експозиції схилу. Фотосинтез пов'язаний з інтенсивністю світла лінійною залежністю до оптимального рівня світлового насичення, за яким слідує зниження інтенсивності унаслідок фотоокислення ферментів. Коефіцієнт використання поглиненої рослиною променистої енергії на фотосинтез не перевищує 10 % за низької освітленості і 1-2% – за високої. Тривалість світлового дня (фотоперіод) відносно постійна на екваторі (близько 12 ч), у вищих широтах змінюється залежно від пори року. Необхідність світла для рослин істотно впливає на структуру угруповань. Розповсюдження водних рослин обмежене поверхневими шарами води. У наземних екосистемах в процесі конкуренції за світло у рослин виробилися певні стратегії, наприклад, швидкий ріст у висоту, використання інших рослин як опори (у ліан), збільшення поверхні листя. У лісах це призводить до ярусної структури фітоценозу. Екологічні групи рослин по відношенню до світла. Рослини діляться на світлові (геліофіти), тіньові (сциофіти, геліофоби), тіньовитривалі (факультативні геліофіти). Геліофіти – види відкритих місць (дуб монгольський, сосна могильна, береза біла, кущисті лишайники, вівсяниця овеча, конюшина повзуча, соняшник та ін.), в сухих місцях зазвичай утворюють розріджений і невисокий покрив. У геліофітів високі витрати на дихання. Характерні ознаки: листя щільне, шкірясте, іноді блискуче з товстою кутикулою, хвоя потовщена, укорочені пагони, опушення, на листах і пагонах сизий восковий наліт – все це захищає лист від перегріву і інтенсивного випаровування. Клітини епідермісу дрібні, паренхіма утворена двома і більше шарами. Співвідношення хлорофілів а: в складає 5:1. Для трав звичайні розеткові форми. Особлива група геліофітів – С4-рослини і САМ-рослини (пустелі, савани). С4-рослини можуть рости навіть при закритих продихах і дуже високій температурі. Світлове насичення фотосинтезу не досягається навіть при найсильнішій освітленості. Виявлено 500 видів покритонасінних з С4-шляхом, який є адаптацією до сухого режиму з високими температурами і інсоляцією. Особливо багато С4- рослин серед родин тонконогові, осокові, портулакові, амарантові, лободові, гвоздикові, молочайні. У САМ-рослин (від Crassulaceae Acid Metabolism) – процес поглинання вуглекислоти відокремлений від синтезу вуглеводнів у часі: вночі СО2 накопичується в органічних кислотах, а вдень включається в послідовність реакцій фотосинтезу при закритих продихах. Сциофіти (тіньові) – не виносять сильного освітлення, ростуть під пологом лісу при сильному затіненні (лісове різнотрав'я, папороті, мохи, кислиця, хвощі, підріст хвойних). Характерні ознаки: ніжне тонке листя з тонкою кутикулою, зазвичай матові, неопушені, пагони витягнуті. Клітини мезофілу великі, паренхіма одношарова, стінки епідермісу тонкі, продихів на одиницю площі менше. Співвідношення хлорофілу а: в менше, ніж у світлолюбних – 3:2. Факультативні геліофіти (тіньовитривалі) займають проміжне положення між двома групами. Легко витримують невелике затінювання. Ефективно використовують бічне розсіяне освітлення, для листя характерне мозаїчне розташування. Це більшість лісових рослин (клени, липи, ліани, багато трав, чагарники). Фотосинтетичний апарат може перебудовуватися при зміні світлового режиму. Так, листя кукурудзи нижнього ярусу, потрапляючи в умови затінювання при сильному розростанні листя середнього і верхнього ярусів, стає тіньовим. У деревних і чагарникових порід тіньова або світлова структура листа часто визначається умовами освітлення попереднього року, коли закладаються бруньки: якщо закладка бруньок йде на світлі, то формується світлова структура, і навпаки. Відношення до світлового режиму змінюється у рослин і в онтогенезі. Проростки і ювенільні рослини багатьох лугових видів і деревних порід більш тіньовитривалі, ніж дорослі особини. Рухи рослин також пов'язані з реакцією на світло: фототропізм, фотонастії. Екологічне значення фототропізму – асимілюючі органи прагнуть зайняти положення, при якому рослина отримуватиме оптимальну кількість світла. Фототропізм спричиняється відтоком ауксинів на затінену сторону. Зміна росту з різних сторін верхівкового пагону призводить до викривлення стебла. У лісі гілки ростуть у напряму відкритого неба. Фотонастії – ріст певних клітин в листі або стеблах під впливом світла. При сильній освітленості сильніше ростуть клітини верхньої поверхні – епінастія, і бічні пагони (шавлія, хризантема, квасоля) або листя в розетках (подорожник, суниця, перстач) приймають горизонтальне положення, в темноті витягуються клітини нижньої поверхні – гіпонастія, і пагін приймає вертикальне положення. Найбільш загальна адаптація рослин до максимального використання ФАР - просторова орієнтація листя. При вертикальному розташуванні листя, як, наприклад, у багатьох злаків, сонячне світло повніше поглинається вранці і ввечері — при нижчому стоянні сонця. При горизонтальній орієнтації листя повніше використовуються промені полуденного сонця. При дифузному розташуванні листя на різних площах сонячна радіація протягом дня використовується найповніше. Зазвичай при цьому листя нижнього ярусу на пагоні відхилене горизонтально, середнього - направлені криво вгору, а верхнього розташовуються майже вертикально. Світло для тварин і для людини має в першу чергу інформаційне значення. Розрізняють види світлолюбні (фотофіли) і тіньолюбні (фотофоби); евріфотні, що виносять широкий діапазон освітленості, і стенофотні, які витримують вузько обмежені умови освітленості. Світло необхідне для орієнтації в просторі. Вже у простих організмів є в клітинах чутливі до світла органели. Бджоли своїм танцем показують шлях польоту до джерела їжі. Встановлено, що фігури танцю (вісімки) співпадають з певним напрямом по відношенню до Сонця. Доведена природжена навігаційна орієнтація птахів: при весняно-осінніх перельотах вони орієнтуються за зірками і Сонцем. У водному середовищі широко поширена біолюмінесценція (риби, головоногі молюски) – здатність світитися для приваблювання здобичі, особин протилежної статі, відлякування ворогів і т. ін. Фототаксис – переміщення у бік найбільшої (позитивний) або найменшої (негативний) освітленості. Так, нічні метелики летять на світло у пошуках партнера, а гримучі змії відчувають інфрачервоне випромінювання. Світло і біоритми. Життя на планеті з моменту виникнення існувало в умовах ритмічних змін середовища. Добова і сезонна зміна комплексу факторів вимагала пристосування до неї всього живого. У процесі еволюції виробилася чітка сумірність і узгодженість біологічних ритмів різних форм життя з періодами циклічних змін комплексу природних умов (і на клітинному, і на біосферному рівні). Світло – головний і постійний первинно-періодичний чинник, що впливає на організми і екосистеми з моменту їх зародження. Зміни світлового режиму найбільш стійкі в своїй динаміці, автономні і не схильні до інших впливів. Виділяють біоритми добові (циркадіанні), сезонні, річні (циркануальні). Циркадіанні (циркадні) ритми – прояв добового ритму, характерний для виду в природних умовах, в умовах незмінної освітленості. Причина -спадково закріплені цикли ендогенних процесів. Наприклад, мімоза на ніч листя складає, на день розпускає – навіть в повній темноті. У птахів і ссавців відомі добові цикли ендокринних залоз і ферментних систем. У арктичних тварин добовий ритм зберігається протягом всього полярного дня, а у інших видів може порушуватися – стерлядь вдень тримається в придонних шарах, вночі плаває скрізь, але якщо цілу добу ясно, то вона так і тримається дна, а якщо темно – вона весь цей час активна. Сезонні ритми. Фізіологічні і біологічні процеси у рослин (процеси репродукції, запасання поживних речовин перед зимовим спокоєм, осіннє забарвлення листя, закладка бруньок та ін.) і більшості видів тварин (шлюбний період, розмноження, линька, сплячка, міграції) виявляються сезонно, з урахуванням зміни пори року. Конкретні погодні умови тільки модифікують протікання цих циклів. Природа цих циклів, як і добових, має еволюційний характер. Циркануальні (цирканні) ритми – це ендогенні біологічні цикли з близько річною періодичністю. Прояв складний, але чітко виражений вплив режиму освітлення. Зокрема, на проходженні онтогенетичних фаз у комах позначається різна тривалість дня. Так, у шовковичного черв'яка Bombyx mori з яєць, відкладених в короткі весняні дні, виводяться самки, яйця яких не впадають в діапаузу, а самки, виведені з яєць довгого літнього дня, відкладають діапаузуючі яйця, забезпечуючи таким чином появу весняного покоління. Таким чином, для рослин світло необхідне в першу чергу, як ресурс для фотосинтезу і транспірації. Для тварин – для інформаційного забезпечення. І для тих і інших – як еволюційний чинник-синхронізатор біологічних ритмів. Волога в житті організмів. Протікання всіх біохімічних процесів в клітинах і нормальне функціонування організму в цілому можливі тільки при достатньому забезпеченні його водою — необхідною умовою життя. Найбільш високий зміст води в періоди активної життєдіяльності (табл. 1) і у молодому віці. Але і в стані спокою рослини не втрачають вологу повністю. В сухих лишайниках міститься до 5-7% води, в зернівках злаків – 12-14%.
Таблиця 1. Вміст води в різних організмах, % від маси тіла
В процесі еволюції у рослин і тварин виробилися численні складні пристосування, що дозволяють підтримувати водний баланс і забезпечувати економне витрачання води. Рослини пустель і степів пристосувалися до гострого дефіциту вологи, болотяні і волого-тропічні рослини – до надлишку, а лісовим видам необхідна висока вологість повітря і помірна вологість ґрунтів. Всі пристосування-адаптації групуються в анатомо-морфологічні, фізіологічні і поведінкові. Джерелами вологи для рослин служать запаси її в ґрунті і атмосфері (опади, тумани, конденсати), для наземних тварин – вода у водоймах, водяна пара в атмосфері і соковита їжа. При аналізі впливу вологи на живі організми важливо враховувати сезонний розподіл і температурний режим місця існування. Різні комбінації змісту води і температури в місці існування створюють безліч різних ситуацій, сприятливих і навпаки. Співвідношення температури і вологості характеризує клімат конкретної території і важливо для вибору популяцією виду стації проживання. Найбільш сприятливі умови складаються в діапазоні температур 17-23°С і в діапазоні відносної вологості повітря 85-100%. За відношенням до вологості розрізняють еврігігробіонтні і стеногігробіонтні організми. Перші живуть в широкому діапазоні вмісту вологи, а для других вона повинна бути або високою, або низькою, або проміжною між першими двома. І серед рослин, і серед тварин є види, які можуть легко переносити дефіцит вологи (копитні, верблюд, варан, миші, лишайники, сукуленти, багато злаків, полини), а можуть і зовсім не виносити посухи (земноводні, плазуни, ластоногі, всі плаваючі рослини, сфагнові мохи, багато папоротей, калужниця та ін.). За способом регуляції водного режиму свого тіла рослини діляться на пойкілогідричні і гомойогідричні. У перших вміст води в тканинах непостійний і залежить від вологозабезпеченості біотопу (наземні водорості, лишайники, мохи, тропічні папороті). У них немає анатомічних пристосувань, що захищають від випаровування, у більшості відсутні продихи. Тіла у них висихають до легко-сухого стану, а у вологу погоду насичуються водою і зеленіють. У інших (більшість покритонасінних рослин) вміст води більш-менш постійний за любої погоди – вони регулюють випаровування шляхом закривання продихів і складання листя, в оболонках клітин у них містяться водонепроникні речовини (кутин, суберин). За відношенням до водного режиму екотопу (екотоп – сукупність факторів місця існування) рослини поділяються на вологолюбні (гідатофіти, гідрофіти, гігрофіти), сухолюбні (ксерофіти) і помірно вологолюбні (мезофіти). Гідатофіти — це водні рослини, цілком або майже цілком занурені у воду. Серед них — квіткові, які повторно перейшли до водного способу життя (елодея, рдесники, водяні жовтці, валіснерія та ін.). Вийняті з води, ці рослини швидко висихають і гинуть. У них редуковані продихи і немає кутикули. Листові пластинки у гідатофітів тонкі, без диференціювання мезофілу, часто розітнуті, що сприяє повнішому використанню послабленого у воді сонячного світла і засвоєнню СО2. Нерідко виражена гетерофілія; у багатьох видів є плаваюче листя, що має світлову структуру. Пагони підтримуються водою, тому часто не мають механічних тканин, в них добре розвинена аеренхима. Коренева система квіткових гідатофітів сильно зредукована, іноді відсутня зовсім, або втратила свої основні функції (у рясок). Поглинання води і мінеральних солей відбувається всією поверхнею тіла. Квітконосні пагоні, як правило, виносять квітки над водою (рідше запилення здійснюється у воді), а після запилення пагоні знову можуть занурюватися, і дозрівання плодів відбувається під водою. Гідрофіти — це рослини наземно-водні, частково занурені у воду, такі, що ростуть по берегах водойм, на болотах. До них можна віднести очерет звичайний, частуху подорожникову, вахту трилисту, калужницу болотну. У них краще, ніж у гідатофітів, розвинені провідні і механічні тканини. Добре виражена аеренхіма. У гідрофітів є епідерміс з продихами, інтенсивність транспірації дуже висока, і вони можуть рости тільки при постійному інтенсивному поглинанні води. Гігрофіти (папороть, росянка, недоторка звичайна, бальзаміни, рис, кислиця) мешкають в дуже вологих місцях і володіють низькою посухостійкістю. У них завжди відкриті продихи і процес транспірації регулюється слабо. Продихи розташовуються з обох боків, нечисленні. Листя крупне тонке. Втрата 15-20% запасу води для них непоправна. Вони ростуть або в глибокій тіні під пологом вологого лісу (тіньові гігрофіти) або на відкритому місці на перезволожених або покритих водою ґрунтах (світлові гігрофіти). Для них характерне товсте слабко розгалужене коріння. В органах велика кількість повітряних порожнин (аеренхіма) для аерації тканин. Мезофіти – здатні нетривало переносити незначні ґрунтову і атмосферну посухи. До них відносяться лугові і багато лісових трав, листяні і хвойні дерева лісів помірної смуги, багато чагарників, більшість сільськогосподарських культур. Продихи розташовані на нижній стороні великого листя. Завдяки регулюванню продихової транспірації, характеризуються великою пластичністю по відношенню до умов зволоження. Для них типові добре розвинені кореневі системи змішаного типу, з густою мережею коріння. Ксерофіти – рослини сухого і жаркого клімату– пустель, степів, саван, в лісовій зоні – рослини сухих сосняків і широколистяних лісів на крутих південних схилах. Вони не виносять перезволоження, але добре пристосувалися до тривалих посух. Для них характерні два способи подолання посухи: активне регулювання водного балансу і здатність виносити сильне висушування тканин. У ксерофітів дуже могутні кореневі системи – за масою в 9-10 разів перевищують надземні органи. Вони або екстенсивного типу (чагарники) – довгі (10-15 м), але мало розгалужені; сягають рівня ґрунтових вод, або інтенсивного (злаки) – охоплюють невеликий об'єм ґрунту (до 1,5 м глибини) і густо гілкуються, максимально засвоюючи вологу. Анатомічні особливості - добре розвинена водопровідна система, мережа жилок на листових пластинках дуже густа, дозволяє швидко поповнювати запаси води, витраченої на транспірацію. Основні риси ксерофітності: дрібні, вузькі, сильно зредуковані листові пластинки, що сприяють зниженню транспірації; зменшення (скидання) листової поверхні в найбільш сухі періоди вегетації (літній листопад); -захист листя від великих втрат вологи на транспірацію завдяки розвитку могутніх покривних тканин, наявність різних виростів, волосків, залозок на епідермісі (едельвейс); - посилений розвиток механічних тканин листа, що запобігають обвисанню при втраті тургору. Види з найбільш вираженими перерахованими властивостями представлені склерофітами (від грец. «склеро» – твердий, жорсткий): саксаул, чортополох, полини, ковила, молочаї та ін. Продихів багато, але вони при нестачі води закриваються. Рослини можуть повністю втрачати все листя і до 15% води. У клітинах склерофітів переважає зв'язана вода. Інша велика група ксерофітів – сукуленти (від латин. «суккулентус» - соковитий, жирний), ростуть в жаркому сухому кліматі там, де проходять короткочасні, але сильні рясні зливи. Під час дощів накопичують в листі (агави, молодило) або стеблах (молочаї, опунція) великі запаси води, а потім поволі її витрачають. Продихів мало, вони дрібні, в поглибленнях, і відкриваються тільки вночі. У північних широтах і високо в горах ростуть аналоги ксерофітів – психрофіти (вологі і холодні місця): мохи, багульник болотний, андромеда і кріофіти (сухі і холодні місця): лишайники, вересові чагарники, арктоус альпійський, брусниця). Вони відчувають нестачу вологи із-за фізіологічної недоступності ґрунтової вологи, обумовленої низькими температурами ґрунтів. Тропофіти – в жарких районах з чергуванням посушливого і вологого сезонів, рослини скидають листя і перебувають в стані глибокого спокою влітку (баобаби в Африці). Еуксерофіти – рослини степів з розетковою і напіврозетковою життєвою формою (Saxifraga omolojensis, S. nivalis, Arenaria sp. – котяча лапка) і сильним опушенням листя. Стіпоксерофіти – теж рослини степових екосистем («стипо» – степ), вузьколистні, дерновинні злаки (вейники, типчаки, тонконіг, міскантус), із сільськогосподарських культур – кукурудза. Вони слабо транспірують, в суху погоду листя згортається в трубочку. Ефемери (весняні і осінні) – однорічні рослини (незабудка піщана, вероніка весняна, мак альпійській), і ефемероїди – багаторічні рослини (крокуси, тюльпани, простріли). Це мешканці посушливих місць існування, які уникають літніх посух у зв'язку з особливостями життєвих циклів. У короткі терміни – за 15-30 днів, рослини встигають пройти весь життєвий цикл і піти на спокій до наступної весни.
Способи регуляції водного балансу у тварин різноманітніше, ніж у рослин. Їх можна розділити на поведінкові, морфологічні і фізіологічні. До поведінкових пристосувань належать пошуки водопоїв, вибір місця існування, риття нор. В норах вологість повітря наближається до 100%, навіть коли на поверхні дуже сухо. Це знижує необхідність випаровування через покриви, економить вологу в організмі. Для пойкілотермних основний шлях збереження водного балансу в пустелі — уникнення зайвих теплових навантажень. До морфологічних способів підтримки нормального водного балансу відносяться утворення, що сприяють затримці води в тілі: раковини наземних равликів, ороговілі покриви рептилій, розвиток епікутикули у комах. Фізіологічні пристосування до регуляції водного обміну — це здатність до утворення метаболічної вологи, економії води при виділенні сечі і калу, розвиток витривалості до зневоднення організму, величина потовиділення і віддача води із слизових оболонок. Серед тварин теж можна виділити три екологічні групи, але із-за рухомого способу життя вони виражені неявно. Гігрофіли – не можуть накопичувати і довго утримувати в тканинах запаси води – багато членистоногих (ногохвістки, комарі, гнус), а також наземні молюски і амфібії. Потребують постійно високої вологості повітря. Мезофіли – тварини, що мешкають в умовах помірної вологості. Їх більшість як серед комах, так і серед ссавців. Ксерофіли – сухолюби і термофіли одночасно, не витримують високу вологість повітря. У них добре розвинені механізми водообміну і функції утримання води в тілі. У плазунів відсутні шкірні залози, виділяється сечова кислота, а не сечовина. У черепахи вода запасається в сечовому міхурі, гризуни воду отримують з їжею. Верблюд, тушканчики, курдючні вівці воду отримують в результаті окислення жирів, при якому утворюється метаболічна вода.
4. Адаптації організмів. При всьому різноманітті форм і механізмів адаптацій живих організмів до дії несприятливих факторів середовища їх можна згрупувати в три основні шляхи: активний, пасивний і уникнення несприятливого впливу. Активний шлях – посилення опірності, розвиток регуляторних здібностей, що дають можливість пройти життєвий цикл і дати потомство, не дивлячись на відхилення умов середовища від оптимальних. Більшою мірою цей шлях властивий гомойотермним організмам, але виявляється і у вищих рослин (прискорення темпів наростання-відмирання пагонів, коріння, швидке цвітіння). Механізми – переважно біохімічні адаптації. Пасивний шлях – підпорядкування життєвих функцій організму зовнішнім умовам. Полягає в економному використанні енергетичних ресурсів при погіршенні умов життя, підвищенні стійкості клітин і тканин. Виявляється в зниженні інтенсивності обмінних процесів, уповільненні швидкості росту і розвитку, літньому листопаді, мінімізації рослин. Найбільш виражений у рослин і пойкілотермних тварин, у ссавців і птахів – тільки у гетеротермних видів. Уникнення несприятливих умов середовища – характерний для всіх живих істот. Проходження життєвих циклів в найбільш сприятливу пору року (активні процеси – у вегетаційний сезон, взимку – стан спокою). Для рослин – захищеність бруньок відновлення і молодих тканин сніжним покривом, підстилкою. Багато дрібних рослин витримують низькі зимові температури, зимуючи під снігом, не маючи ніяких адаптивних рис у вигляді зміни органів або клітин. Стани спокою. Всі рослини і тварини готуються до зими, при цьому сповільнюються фізіолого-біохімічні процеси. У рослин припиняється ріст. У деревних порід, що впадають в стан спокою, інтенсивність дихання знижується до 1/200 – 1/400 від літньої. Органічний спокій характерний для плодів, бульб, бруньок (сплячі бруньки). Вони не проростають, не розпускаються до весни або до пори, поки не отримають «сигнал» чинників середовища, поки не відбудуться біохімічні реакції в ембріональних клітинах і тканинах. Глибокий спокій – наступає одночасно з органічним; ступінь глибини залежить від виду рослин і умов осені; забезпечує морозостійкість. Вимушений спокій – затримка весняного розвитку із-за несприятливих умов. У тварин теж декілька станів спокою. Сплячка – літня – через високі температури і дефіцит води, зимова – через холод. Не завжди у ссавців під час зимового сну сповільнюються обмінні процеси – у бурих і білих ведмедів взимку народжуються дитинчата. Анабіоз – стан організму, при якому життєві процеси настільки завмирають, що ознаки життя відсутні. Організм зневоднюється і тому може переносити дуже низькі температури (до -271°С), але при цьому не відбувається порушення макромолекул в клітинах. Анабіоз характерний для спор, насіння, висохлих лишайників, простих одноклітинних. Діапауза – стан тимчасової зниженої фізіологічної активності – властива членистоногим. У цей період сповільнюються процеси обміну, підвищується стійкість до несприятливих умов середовища. Розрізняють зимову і літню (у дощових черв'яків, у кільчастого шовкопряда, дубової і горіхової павлиноочки, листоїдів) діапаузи. Може наступати на певній стадії розвитку (лялечки, гусениці, яйця), триває від декількох тижнів до року і часто не пов'язана з погіршенням умов. У сумчастих може непередбачувано затриматися розвиток плоду до настання сприятливого поєднання факторів середовища – у відповідь «сплячка».
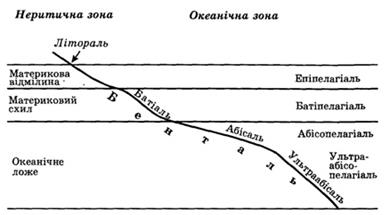
. Лекція 4. ОСНОВНІ СЕРЕДОВИЩА МЕШКАННЯ Водне середовище мешкання. Водне середовище мешкання Вода покриває 71% площі земної поверхні. Основна маса води зосереджена в морях і океанах – 94-98%, в полярних льодах міститься близько 1,2% води і зовсім мала частка – менше 0,5% зосереджена в прісних водах річок, озер і боліт. У водному середовищі мешкає близько 150 000 видів тварин і 10 000 рослин, що складає відповідно всього 7 і 8 % від загального числа видів на Землі. У морях та океанах, як в горах, виражена вертикальна зональність. Особливо сильно різняться за екологічними умовами пелагіаль – вся товща води та бенталь – дно. Пелагіаль по вертикалі ділиться на зони: епіпелагіаль, батіпелагіаль, абісопелагіаль ультраабісопелагіаль (рис. 3).
Рис. 3 Екологічні зони Світового океану
Залежно від крутизни спуску і глибини на дні виділяють декілька зон, яким відповідають зони пелагіалі: - літоральна – кромка берега, що заливається під час припливів; - супраліторальна – частина берега вища за верхню припливну межу, куди долітають бризки прибою; - субліторальна – плавне пониження суші до 200м; - батіальна – круте пониження суші (материковий схил); - абісальна – плавне пониження дна океанського ложа; глибина обох зон разом досягає 3-6 км. - ультраабісальна – глибоководні западини від 6 до 10 км. Екологічні групи гідробіонтів. Найбільшою різноманітністю життя відрізняються теплі моря і океани (40000 видів тварин) в області екватора і тропіках, на північ і південь відбувається збіднення флори і фауни морів в сотні разів. Що стосується розподілу організмів безпосередньо в морі, то основна маса їх зосереджена в поверхневих шарах (эпіпелагіаль) і в субліторальній зоні. Залежно від способу пересування і перебування в певних шарах, морські мешканці підрозділяються на три екологічні групи: нектон, планктон і бентос. Нектон (nektos – плаваючий) - крупні тварини, що активно пересуваються, здатні долати великі відстані і сильні течії: риби, кальмари, ластоногі, кити. У прісних водоймах до нектону відносяться земноводні і безліч комах. Планктон (planktos – блукаючий) – сукупність рослин (фітопланктон: діатомові, зелені і синьо-зелені водорості) і дрібних тваринних організмів (зоопланктон: джгутиконосці, дрібні ракоподібні, крилоногі молюски, медузи, гребнівики, деякі черв'яки), що мешкають на різній глибині, але не здатні до активних пересувань і до протистояння течіям. До складу планктону входять і личинки тварин, утворюючи особливу групу – нейстон. Це пасивно плаваюче «тимчасове» населення самого верхнього шару води, представлене різними тваринами (десятиногі, вусоногі і веслоногі ракоподібні, голкокожі, поліхети, риби, молюски) в личинковій стадії. Личинки, дорослішаючи, переходять в нижні шари пелагіалі. Вище нейстона розташовується плейстон – це організми, у яких верхня частина тіла росте над водою, а нижня – у воді (ряска – Lemma, сифонофорита ін.). Планктон відіграє важливу роль в трофічних зв'язках біосфери, оскільки є їжею для багатьох водних мешканців, зокрема основним кормом для вусатих китів (Myatcoceti). Бентос (benthos – глибина) – гідробіонти дна. Представлений в основному прикріпленими або такими, що поволі пересуваються, тваринами (зообентос: форамініфери, риби, губки, кишковопорожнинні, черв'яки, молюски, асцидії), більш чисельними на мілководді. На мілководді в бентос входять і рослини (фітобентос: діатомові, зелені, бурі, червоні водорості, бактерії). На глибині, де немає світла, фітобентос відсутній. Найбільш багаті на фітобентос кам'янисті ділянки дна. В озерах зообентос менш рясний і різноманітний, ніж в морі. Його утворюють прості (інфузорії, дафнії), п'явки, молюски, личинки комах та ін. Фітобентос озер утворений вільно плаваючими діатомеями, зеленими і синьо-зеленими водоростями; бурі та червоні водорості відсутні.
Фізико-хімічна характеристика водного середовища. Висока щільність водного середовища визначає особливий склад і характер зміни факторів життєзабезпечення. Одні з них такі ж, що і на суші – тепло, світло, інші специфічні: тиск води (з глибиною збільшується на 1 атмосферу на кожні 10 м), вміст кисню, склад солей, кислотність. Завдяки високій щільності середовища, значення тепла і світла з градієнтом висоти змінюються набагато швидше, ніж на суші. Тепловий режим. Для водного середовища характерний менший прихід тепла, тому що значна частина його відбивається і не менш значна частина витрачається на випаровування. У порівнянні з наземним середовищем вода має меншу амплітуду коливань добових і сезонних температур. Більше того, водойми істотно вирівнюють хід температур в атмосфері прибережних районів. При відсутності льодового панцира моря в холодну пору року водойми мають утеплюючий вплив на прилеглі території суші, а влітку – охолоджувальний і зволожуючий. Діапазон значень температури води в Світовому океані складає 38° (від -2 до +36°С), в прісних водоймах – 26° (від -0,9 до +25°С). З глибиною температура води різко падає. До 50 м спостерігаються добові коливання температури, до 400 м – сезонні, глибше вона стає постійною, +1- +3°С. Оскільки температурний режим у водоймах порівняно стабільний, їх мешканцям властива стенотермність. У зв'язку з різним ступенем прогрівання верхніх і нижніх шарів впродовж року, припливами і відливами, течіями, штормами відбувається постійне перемішування водних шарів. Роль перемішування води для водних мешканців виключно велика, оскільки при цьому вирівнюється розподіл кисню і поживних речовин усередині водойм, забезпечуючи обмінні процеси між організмами і середовищем. У стоячих водоймах (озерах) помірних широт навесні і осінню має місце вертикальне перемішування, і в ці сезони температура у всьому водоймі стає однорідною, тобто наступає гомотермія. Влітку і зимою в результаті різкого посилення прогрівання або охолодження верхніх шарів перемішування води припиняється. Це явище називається температурною дихотомією, а період тимчасового застою – стагнацією (літньою або зимовою). Влітку легші теплі шари залишаються на поверхні, розташовуючись над важкими холодними. Взимку, навпаки, в придонному шарі тепліша вода, оскільки безпосередньо під льодом температура поверхневих вод менша +4°С і вони через фізико-хімічні властивості води стають більш легкими, ніж вода з температурою, вищою за +4°С. У періоди стагнацій чітко виділяються три шари: верхній (епілімніон) з найбільш різкими сезонними коливаннями температури води, середній (металімніон або термоклін), в якому відбувається різкий стрибок температур, і придонний (гіполімніон), в якому температура протягом року змінюється слабо. У періоди стагнацій в товщі води утворюється дефіцит кисню – влітку в придонній частині, а взимку і у верхній, внаслідок чого відбуваються замори риби. Світловий режим. Інтенсивність світла у воді сильно послаблена через його віддзеркалення поверхнею і поглинання самою водою. Це сильно позначається на розвитку фотосинтезуючих рослин. Поглинання світла тим сильніше, чим менше прозорість води, яка залежить від кількості зважених в ній частинок (мінеральні суспензії, планктон). Зменшується вона при бурхливому розвитку дрібних організмів влітку, а в помірних і північних широтах – ще і взимку, після встановлення льодового покриву і укриття його зверху снігом. Прозорість характеризують граничною глибиною, на якій ще видно білий диск, що спеціально опускається, діаметром близько 20 см (диск Секі). Найпрозоріші води — в Саргасовому морі: диск видно до глибини 66,5 м. У Тихому океані диск Секкі видно до 59 м, в Індійському — до 50 м, в дрібних морях — до 5—15м. Прозорість річок в середньому 1—1,5 м, а в найкаламутніших річках всього декілька сантиметрів. У океанах, де вода дуже прозора, на глибину 140 м проникає 1% світловій радіації, а в невеликих озерах на глибину 2 м проникають всього лише десят
|
||||||||||||
|
|
Последнее изменение этой страницы: 2021-07-19; просмотров: 171; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.188.69.167 (0.032 с.) |