Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Экологические основы природопользованияСодержание книги
Поиск на нашем сайте
Экологические основы природопользования
Лекция 3. Взаимосвязи живого и косного вещества на уровне «организм» ↔ «среда обитания» Законы взаимодействия организмов и абиотических факторов. Модель «Колесо жизни» (рис. 2а, б) более всего подходит для объяснения двух основных законов взаимодействия организмов и абиотических факторов: это закон оптимума и закон ограничивающих факторов (минимума Либиха). Очень кратко, с помощью Б.Небела, опишем их суть. «Для разных видов, условия в которых они особенно хорошо себя чувствуют, неодинаковы. Например, некоторые растения предпочитают очень влажную почву, другие – относительно сухую. Одни требуют сильной жары, другие лучше переносят более холодную среду и т.д. В лабораторных экспериментах эти различия проявляются особенно четко. Если провести эксперимент, где контролируются все абиотические факторы, и только один из них, например, температуру, можно изменять, то результаты показывают, что по мере повышения температуры от некой точки, при которой рост вообще невозможен, растение развивается всё лучше и лучше, пока интенсивность роста не достигнет максимального уровня. Затем при дальнейшем повышении температуры растение будет чувствовать себя хуже и хуже, у него появятся нарушения и, в конечном итоге, оно погибнет. Графически это выражается всем известной параболой в системе координат, на оси ординат которой откладывается скорость роста, а на оси абсцисс – температура. Полученная парабола роста четко делится на зоны – зону оптимума, т.е. такой диапазон температур, при котором скорость роста максимальна; зону устойчивости – это интервал температур, от минимального до максимального значения, при котором рост вообще возможен; максимальные и минимальные температуры, пригодные для жизни, - это пределы устойчивости. Между зоной оптимума и пределами устойчивости по мере приближения к последним, растение испытывает всё нарастающий стресс – это стрессовые зоны в рамках диапазона устойчивости. Формулировка этого фундаментального закона: Для каждого вида (растений и животных) существует оптимум, стрессовые зоны и пределы устойчивости в отношении каждого средового фактора» (конец цит.). Обратите внимание на модель «Колесо жизни» - это ничто иное, как состояние организма под оптимальным воздействием каждого фактора. Но если какой-то один из них будет в недостатке, т.е. в колесе окажется «дырка» (рис. 2 б), то это состояние описывается законом лимитирующих факторов: «Даже единственный фактор за пределами зоны своего оптимума приводит к стрессовому состоянию организма и в пределе – к его гибели. Такой фактор называют лимитирующим» (Реймерс, Начала экологических знаний). По этим и другим законам есть довольно обширная литература, поэтому отсылаю читателей к работам Ю.Одума, Н.Ф.Реймерса, Н.М.Черновой и др. В электронном приложении к этому пособию даны механизмы и характеристики воздействия важнейших абиотических факторов, обозначенных в модели. Характеристика абиотических факторов и закономерностей их воздействия предельно логично объясняет принципы экологической классификации. Различают три основных типа экологических пирамид: пирамида чисел (численности), пирамида биомассы и пирамиды энергии. 1) пирамиды чисел, основанные на подсчете организмов каждого трофического уровня; 2) пирамиды биомассы, в которых используется суммарная масса (обычно сухая) организмов на каждом трофическом уровне; 3) пирамиды энергии, учитывающие энергоемкость организмов каждого трофического уровня. Пирамиды энергии считаются самыми важными, поскольку они непосредственно обращаются к основе пищевых отношений — потоку энергии, необходимой для жизнедеятельности любых организмов. Пирамида биомассы Пирамида биомассы отражает более полно пищевые взаимоотношения в экосистеме, так как в ней учитывается суммарная масса организмов (биомасса) каждого трофического уровня. Прямоугольники в пирамидах биомассы отображают массу организмов каждого трофического уровня, отнесенную к единице площади или объема. Пирамиды биомассы, так же, как и пирамиды численности, могут быть не только правильной формы, но и перевернутыми (обращенными) рис.8.
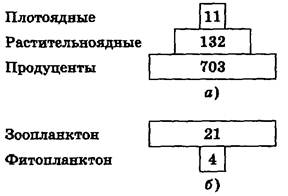
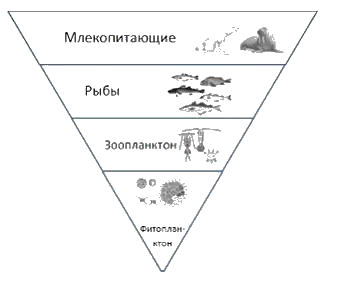
Рис. 7.5. Пирамиды биомасс биоценозов кораллового рифа (а) и пролива Ла-Манш (б).Цифры означают биомассу в граммах сухого вещества,приходящегося на 1 кв.м Для большинства наземных экосистем (луг, поле и др.) суммарная биомасса каждого последующего трофического уровня пищевой цепи уменьшается. Это создает пирамиду биомасс, где существенно преобладают продуценты, а над ними располагаются постепенно уменьшающиеся трофические уровни консументов, т.е. пирамида биомасс имеет правильную форму. Например: · в среднем из 1000 кг растений образуется 100 кг тела растительноядных животных – консументов первого порядка (фитофагов). Плотоядные животные – консументы второго порядка, поедающие растительноядных, могут синтезировать из этого количества 10 кг своей биомассы. А хищники – консументы третьего порядка, питающиеся плотоядными животными, синтезируют только 1 кг своей биомассы. В водных экосистемах (озеро, пруд и др.) пирамида биомасс может быть перевернутой, где биомасса консументов преобладает над биомассой продуцентов. Это объясняется тем, что в водных экосистемах продуцентом является микроскопический фитопланктон, быстро растущий и размножающийся), который в достаточном количестве непрерывно поставляет живую пищу консументам, намного медленно растущим и размножающимся. Зоопланктон (или другие животные, питающиеся фитопланктоном) накапливают биомассу годами и десятилетиями, тогда как фитопланктон имеет крайне короткий период жизни (несколько дней или часов).
Числа на рисунке относятся к таким показателям, а стрелки символизируют поток энергии и поступление веществ. Пирамида энергий отражает динамику прохождения массы пищи через пищевую (трофическую) цепь, что принципиально отличает ее от пирамид численности и биомасс, отражающих статику системы (количество организмов в данный момент). На форму этой пирамиды не влияют изменения размеров и интенсивности метаболизма особей. Если учтены все источники энергии, то пирамида всегда будет иметь типичный вид (в виде пирамиды вершиной вверх), согласно второму закону термодинамики.
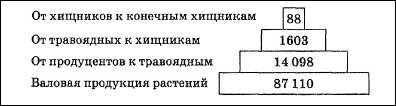
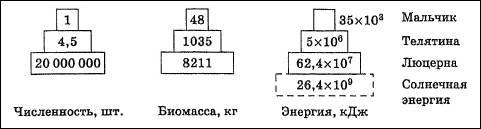
Рис. 7. Пирамида энергии: цифры - количество энергии, кДж * м-2 *r-1 Пирамиды энергий позволяют не только сравнивать различные биоценозы, но и выявлять относительную значимость популяций в пределах одного сообщества. Они являются наиболее полезными из трех типов экологических пирамид, однако получить данные для их построения труднее всего. Одним из наиболее удачных и наглядных примеров классических экологических пирамид служат пирамиды, изображенные на рис. 8 Николайкин Н. И. Экология: Учеб. для вузов / Н. И. Николайкин, Н. Е. Николайкина, О. П. Мелехова. - 3-е изд., стереотип. - М.: Дрофа, 2004.. Они иллюстрируют условный биоценоз, предложенный американским экологом Ю. Одумом. «Биоценоз» состоит из мальчика, питающегося только телятиной, и телят, которые едят исключительно люцерну.
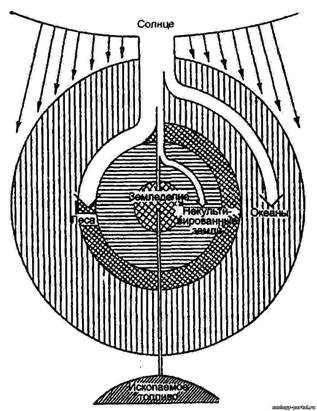
Рис. 8. Экологические пирамиды (по Ю. Одуму). Без соблюдения масштаба Согласно закону экологической пирамиды энергий, или правилу десяти процентов (его называют также законом, или правилом Р. Линдемана, по имени сформулировавшего его в 1942 г. немецкого ученого): «с одного трофического уровня экологической пирамиды на другой в среднем переходит около 10% поступающей на предыдущий уровень энергии». На самом деле потеря бывает не десятикратной, а несколько меньше, а в других случаях – больше, но порядок чисел сохраняется». На практике это означает, что довольно с большой точностью можно спрогнозировать, сколько телят сможет прокормить пастбище известной площади с известной растительной биомассой, каких привесов могут достигнуть эти телята и сколько мяса можно будет выставить на продажу в конце года. Так чисто экологические проблемы легко и быстро переходят в проблемы экономические. Биомасса в наземных экосистемах убывает от продуцентов к редуцентам. Согласно правилу 10% убывает и биогеохимическая энергия. Обратных энергетических пирамид, как и двигателей второго рода, не бывает. Но об обратных пирамидах приходится говорить, когда в пищевую цепь постоянно попадают вредные вещества. Их накопление происходит очень интенсивно, это иллюстрируется примером с пестицидом ДДТ (дихлордифенилтрихлорэтан), относящимся к группе дустов*. ________________________________________________________________________ *Токсикологические свойства и характеристики ЛД 50 для различных экспериментальных животных при введении ДДТ через рот составляет 250-400 мг/кг. Установлено, что ДДТ способен накапливаться в жировой ткани у животных и человека. Несмотря на его низкую растворимость в воде, в водных организмах происходит его биоконцентрация, и содержание ДДТ в ряде гидробионтов доходит до десятков миллиграмм на 1 кг живой массы. [5] Интересно отметить, что в присутствии барбитуратов (люминала) метаболизм ДДТ в организме человека и животных протекает значительно быстрее, и препараты за сравнительно короткий срок выводятся из организма. [5] ДДТ обладает устойчивостью к разложению. Ни свет, ни ферменты, ни критичные температуры не способны сильно повлиять на процесс разложения ДДТ. В итоге, при попадании в окружающую среду, ДДТ проникает в пищевые цепи. В них токсикант накапливается в значительных количествах: сначала в растениях, потом в животных организмах и, в итоге, в человеческом теле. Расчеты исследователей показали, что на каждом звене пищевой цепи происходит рост содержания ДДТ в десять раз: · Ил, содержащий ДДТ – 1х · Растения (водоросли) – 10х · Мелкие организмы (рачки) – 100х · Рыбы – 1000х · Хищные рыбы – 10000х Низкая растворимость в воде и высокая растворимость в жирах и обусловливает задержку ДДТ в жировых клетках. Скорость накопления вещества в организме варьирует в зависимости от концентрации, длительности воздействия, вида живого объекта и условий окружающей среды. Высокая степень удержания ДДТ говорит о том, что токсические эффекты способны проявляться в течение большого промежутка времени, а также на значительном географическом удалении от места воздействия. Организмы высоких пищевых уровней имеют тенденцию к накоплению больших количеств ДДТ в сравнении с организмами низших пищевых уровней. В организмах мигрирующих животных ДДТ способен транспортироваться по всему миру, а также воздушным и океаническим потоками. [8] _____________________________________________________________________________ Н.Ф. Реймерс в книге «Начала экологических знаний» приводит следующее правило накопления вредных веществ: «Если полезное при его излишке легко выводится из организма, то вредное не только плохо выводится, но и накапливается в пищевой цепи» (конец цитаты). Отсюда мы заключаем, что между «вредным» и «полезным» существует принципиальное отличие, которое заключается, прежде всего, в искусственном происхождении «вредного» и природном происхождении «полезного». В живой природе на каждую сложную молекулу есть свой «упроститель» - чаще всего это фермент или группа ферментов. Но вся беда в том, человек, синтезируя все новые и новые химические соединения, совершенно не задумывается о способах их превращения в простые молекулы - продукты любого естественного круговорота. Нагроможденные друг на друга атомы синтетических веществ накапливаются в организмах так стремительно, как это показано на примере с ДДТ. Накопление идет как в клетках, так и в межклеточных пространствах, при этом механизмы вывода существуют только для «своих» молекул, а чужеродные, неспособные сами разлагаться до простых веществ, молекулы «болтаются» по организму, безмерно его засоряя. Причем, периоды полураспада этого и похожих пестицидов составляют от 10 до 20 лет. Есть искусственно созданные, сравнительно простые химические соединения, например, витамины, которые, как нам кажется, «усваиваются» организмом. Но, учитывая принцип Луи Пастера о киральной чистоте основных жизненно важных молекул (сахара – правые, аминокислоты – левые), мы должны помнить, что промышленно синтезированные подобные вещества образуют только рацемические смеси, такие же, которые существуют в неживой природе. Сколько бы человек не старался синтезировать только левые или только правые формы любых молекул, получается снова и снова соотношение 50:50. Значит, всё, что не имеет натурального происхождения, полезно для человека ровно на 50%, если это лекарственные или пищевые продукты. А если речь идет о моющих средствах или пестицидах, которые не разлагаются на простые вещества, то такие химические соединения вообще не имеют права на существование, поскольку не могут встраиваться в природные биогеохимические циклы. Вместе с тем, если разработан механизм их редуцирования до простых соединений, пусть даже до рацемических смесей, такие соединения могут почти безболезненно использоваться человеком. Продуктивность экосистем Продуктивность экосистем тесно связана с потоком энергии, проходящим через ту или иную экосистему. В каждой экосистеме часть приходящей энергии, попадающей в трофическую сеть, накапливается в виде органических соединений. Безостановочное производство биомассы (живой материи) — один из фундаментальных процессов биосферы. Органическое вещество, создаваемое продуцентами в процессе фотосинтеза или хемосинтеза, называют первичной продукцией экосистемы (сообщества). Количественно ее выражают в сырой или сухой массе растений или в энергетических единицах —эквивалентном числе ккалорий или джоулей. Первичной продукцией определяется общий поток энергии через биотический компонент экосистемы, а следовательно, и биомасса живых организмов, которые могут существовать в экосистеме (рис. 12.33).
Рис. 12.33. Первичная продукция больших подразделений биосферы (из Ф. Рамада, 1981) Примечание: интенсивность продукции пропорциональна густоте штриховки __________________________________________________________________ Теоретически возможная скорость создания первичной биологической продукции определяется возможностями фотосинтетического аппарата растений. А как известно, лишь часть энергии света, получаемой зеленой поверхностью, может быть использована растениями. Из коротковолнового излучения Солнца только 44% относится к фотосинтетически активной радиации (ФАР) — свет по длине волны, пригодный для фотосинтеза. Максимально достигаемый в природе КПД фотосинтеза 10—12% энергии ФАР, что составляет около половины от теоретически возможного, отмечается в зарослях джугары и тростника в Таджикистане в кратковременные, наиболее благоприятные периоды. КПД фотосинтеза в 5% считается очень высоким для фитоценоза. В целом по земному шару усвоение растениями солнечной энергии не превышает 0,1 % из-за ограничения фотосинтетической активности растений множеством факторов, среди них таких, как недостаток тепла и влаги, неблагоприятные физические и химические свойства почвы и т. д. Средний коэффициент использования энергии ФАР для территории России равен 0,8%, на европейской части страны составляет 1,0—1,2%, а в восточных районах, где условия увлажнения менее благоприятны, не превышает 0,4— 0,8%. Скорость, с которой растения накапливают химическую энергию, называют валовой первичной продуктивностью (ВПП). Около 20% этой энергии расходуется растениями и?1| дыхание и фотодыхание. Скорость накопления органического веще4| ства за вычетом этого расхода называется чистой первичной иро-| дуктивностью (ЧПП). Это энергия, которую могут использовать| организмы следующих трофических уровней. Количество органического вещества, накопленного гетеротрофными организмами, называется вторичной продукцией. Вторичную продукцию вычисляют отдельно для каждого трофического уровня, так как прирост массы на каждом из них происходит за счет энергии, поступающей с предыдущего. Гетеротрофы, включаясь в трофические цепи, в конечном итоге живут за счет чистой первичной продукции сообщества. Полнота ее расхода в разных экосистемах различна. Постеленное увеличение общей биомассы продуцентов отмечается, если скорость изъятия первичной продукции в цепях питания отстает от темпов прироста растений. Мировое распределение первичной биологической продукции весьма неравномерно. Чистая продукция меняется от 3000 г/м2/год до нуля в экстрааридных пустынях, лишенных растений, или в условиях Антарктиды с ее вечными льдами на поверхности суши, а запас биомассы — соответственно от 60 кг/м2 до нуля. Р. Уиттекер (1980) делит по продуктивности все сообщества на четыре класса. 1. Сообщества высшей продуктивности, 3000—2000 г/м2/год. Сюда относятся тропические леса, посевы риса и сахарного тростника. Запас биомассы в этом классе продуктивности весьма различен и превышает 50 кг/м2 в лесных сообществах и равен продуктивности у однолетних сельскохозяйственных культур. 2. Сообщества высокой продуктивности, 2000—1000 г/м2/год. В этот класс включены листопадные леса умеренной полосы, луга при применении удобрений, посевы кукурузы. Максимальная биомасса приближается к биомассе первого класса. Минимальная биомасса соответственно равна чистой биологической продукции однолетних культур. 3. Сообщества умеренной продуктивности, 1000—250 г/м2/год. К этому классу относится основная масса возделываемых сельскохозяйственных культур, кустарники, степи. Биомасса степей меняется в пределах 0,2—5 кг/м2. 4. Сообщества низкой продуктивности, ниже 250 г/м^год — пустыни, полупустыни (в отечественной литературе их называют чаще опустыненными степями), тундры. Биомасса и первичная продуктивность основных типов экосистем представлена в табл. 12.6. Таблица 12.6 Как проявляется аллелопатия Воздействие выделяемых химических веществ может быть трех типов: 1. Отрицательное – проявляется в угнетении роста и развития соседних растений, снижении способности к зимовке, низкой урожайности, гибели культур. Выделяемые вещества накапливаются почвой и постепенно создают аллелопатическую среду, которая благоприятно влияет на одни растения, и угнетает другие;
2. Положительное – проявляется в повышенной вегетации соседних культур, высокой урожайности и устойчивости растений к вредным факторам среды. Происходит это за счет того, что некоторые растения выделяют вещества, угнетающие рост сорняков, а также препятствующие распространению вредителей и различных микробных и грибковых заболеваний;
3. Нейтральное – данный вид взаимодействия никак не сказывается на росте и развитие соседствующих растений.
Примеры аллелопатии растений Примеры несовместимости культур: Вишня несовместима с такими культурами, как яблоня, груша, абрикос; Бобы с луком, чесноком, сельдереем; Кабачки с помидорами; Капуста с томатами, клубникой и фасолью; Картофель с малиной, тыквой, огурцами, помидорами, подсолнечником, вишней, яблоней; Морковь с сельдереем, свеклой, хреном; Перец с бобовыми, огурцами, сельдереем и фенхелем; Слива с деревьями груши, яблони; Томаты с капустой кольраби, бобовыми, огурцами; Лук с бобовыми и капустой; Редис и редька с бобовыми и земляникой. Таким образом, если правильно распределить различные культуры на участке, с учетом аллелопатического влияния, то возможно предотвратить угнетение и плохой рост некоторых растений, добиться высокой урожайности, при этом избавится от вредителей и болезней.
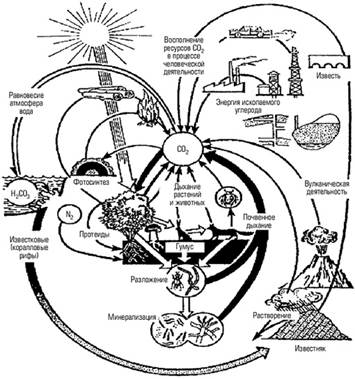
Барри Коммонер сказал, что “природа знает лучше”. Она не только знает, но и подсказывает человеку: “Смотри, что будет с тобой, если ты поддашься вредным соблазнам...” Известный наш лесовод опирался на учение В. В. Докучаева и работы К. Мебиуса — автора понятия биоценоза. Современная формулировка правила взаимоприспособленности организмов в биоценозе Д. Мебиуса — Г. Ф. Морозова: Круговорот азота в биосфере В атмосфере и живом веществе содержится менее 2% всего азота на Земле, но именно он поддерживает жизнь на планете. Азот входит в состав важнейших органических молекул — ДНК, белков, липопротеидов, АТФ, хлорофилла и др. В растительных тканях его соотношение с углеродом составляет в среднем 1: 30, а в морских водорослях I: 6. Биологический цикл азота поэтому также тесно связан с углеродным. Молекулярный азот атмосферы недоступен растениям, которые могут усваивать этот элемент только в виде ионов аммония, нитратов или из почвенных или водных растворов. Поэтому недостаток азота часто является фактором, лимитирующим первичную продукцию — работу организмов, связанную с созданием органических веществ из неорганических. Тем не менее атмосферный азот широко вовлекается в биологический круговорот благодаря деятельности особых бактерий (азотфиксаторов). В круговороте азота большое участие также принимают аммонифицирующие микроорганизмы. Они разлагают белки и другие содержащие азот органические вещества до аммиака. В аммонийной форме азот частью вновь поглощается корнями растений, а частью перехватывается нитрифицирующими микроорганизмами, что противоположно функциям группы микроорганизмов — денитрификаторов.
Рис. 8. Круговорот азота В анаэробных условиях в почвах или водах они используют кислород нитратов для окисления органических веществ, получая энергию для своей жизнедеятельности. Азот при этом восстанавливается до молекулярного. Азотфиксация и денитрификация в природе приблизительно уравновешены. Цикл азота, таким образом, зависит преимущественно от деятельности бактерий, тогда как растения встраиваются в него, используя промежуточные продукты этого цикла и намного увеличивая масштабы циркуляции азота в биосфере за счет продуцирования биомассы. Роль бактерий в круговороте азота настолько велика, что если уничтожить только 20 их видов, жизнь на нашей планете прекратится. Небиологическая фиксация азота и поступление в почвы его окислов и аммиака происходит также с дождевыми осадками при ионизации атмосферы и грозовых разрядах. Современная промышленность удобрений фиксирует азот атмосферы в размерах, превышающих природную фиксацию азота, в целях увеличения продукции сельскохозяйственных растений. В настоящее время деятельность человека все сильнее влияет на круговорот азота, в основном в сторону превышения перевода его в связанные формы над процессами возврата в молекулярное состояние.
Круговорот серы в биосфере
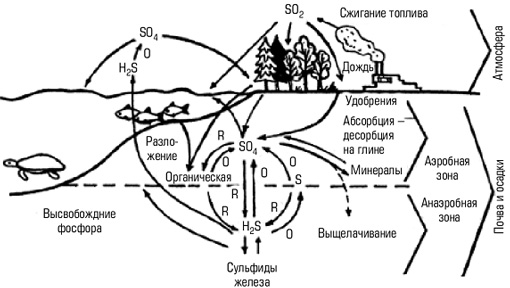
Круговорот серы, необходимой для построения ряда аминокислот, отвечает за трехмерную структуру белков, поддерживается в биосфере широким спектром бактерий. В отдельных звеньях этого цикла участвуют аэробные микроорганизмы, окисляющие серу органических остатков до сульфатов, а также анаэробные редукторы сульфата, восстанавливающие сульфаты до сероводорода. Кроме перечисленных группы серобактерий окисляют сероводород до элементарной серы и далее до сульфатов. Растения усваивают из почвы и воды только ионы SO2-4. Кольцо в центре иллюстрирует процесс окисления (О) и восстановления (R), благодаря которым происходит обмен серы между фондом доступного сульфата и фондом сульфидов железа, находящимся глубоко в почве и осадках.
Рис. 10. Круговорот серы. Кольцо в центре иллюстрирует процесс окисления (0) и восстановления (R), благодаря которым происходит обмен серы между фондом доступного сульфата и фондом сульфидов железа, находящимся глубоко в почве и осадках Основное накопление серы происходит в океане, куда ионы сульфатов непрерывно поступают с суши с речным стоком. При выделении из вод сероводорода сера частично возвращается в атмосферу, где окисляется до диоксида, превращаясь в дождевой воде в серную кислоту. Промышленное использование большого количества сульфатов и элементарной серы и сжигание горючих ископаемых поставляют в атмосферу большие объемы диоксида серы. Это вредит растительности, животным, людям и служит источником кислотных дождей, усугубляющих отрицательные эффекты вмешательства человека в круговорот серы.
Описание Моря теряют из-за испарения больше воды, чем получают с осадками, на суше — положение обратное. Вода непрерывно циркулирует на земном шаре, при этом её общее количество остаётся неизменным. Три четверти поверхности земного шара покрыты водой. Водную оболочку Земли называют гидросферой. Большую её часть составляет соленая вода морей и океанов, а меньшую — пресная вода озер, рек, ледников, грунтовые воды и водяной пар. На земле вода существует в трех агрегатных состояниях: жидком, твердом и газообразном. Без воды невозможно существование организмов. В любом организме вода является средой, в которой происходят химические реакции, без которых не могут жить организмы. Вода является самым ценным и самым необходимым веществом для жизнедеятельности живых организмов. Постоянный обмен влагой между гидросферой, атмосферой и земной поверхностью, состоящий из процессов испарения, передвижения водяного пара в атмосфере, его конденсации в атмосфере, выпадения осадков и стока, получил название круговорота воды в природе. Атмосферные осадки частично испаряются, частично образуют временные и постоянные водоемы, частично — просачиваются в землю и образуют подземные воды. Круговорот воды приводится в движение энергией Солнца. Солнце нагревает воду в океанах и морях, и она испаряется, преобразуясь в водяной пар. Параллельный процесс происходит и на суше: вода испаряется с нагретой Солнцем поверхности Земли или испаряется растениями в результате транспирации. В процессе адвекции водяной пар перемещается с воздушными массами, пока, в конце концов, не оказывается в зоне с низкой температурой. Это вызываетконденсацию влаги в облаках. Облака продолжают перемещаться вместе с воздухом, в то время как сконденсированные капельки воды в них перемешиваются, слипаются и растут в размерах. В итоге вода выпадает в виде осадков над сушей или океаном; при этом океан испаряет больше влаги в атмосферу, чем приобретает от осадков, а суша — наоборот, получает с осадками больше, чем с неё испаряется. Некоторые осадки выпадают в виде снега или града, дождя со снегом, и могут накапливаться в ледяных шапках и ледниках, которые хранят замороженную воду в течение от нескольких месяцев до десятков тысяч лет. Но даже в таком виде незначительный обмен льдов с атмосферой сохраняется: действует сублимация. В то время, когда температура в зоне отложений повышается, начинается таяние, и вода активно исходит из этих источников. Большая же часть воды возвращается из атмосферы в виде дождя. Часть выпавших осадков перехватывается листвой растений, не достигнув почвы. Попав на сушу, вода перетекает по земле в виде рек, двигаясь к океанам. Часть из этой воды впитывается в грунт в результате инфильтрации, проникает глубоко в землю и пополняет водоносные горизонты грунтовых вод, которые также аккумулируют в себе пресную воду в течение длительного времени. Под землёй, как и на ей поверхности, тоже существует движение водяных масс, и вода движется, меняя своё местоположение. Грунтовые воды обмениваются водой с поверхностью в виде родников и артезианских скважин (разгрузка грунтовых вод). Эта, а также небольшая часть впитавшейся в землю, но не достигшей уровня водоносных горизонтов воды, попадает назад в поверхностные водные объекты и океан. Доля воды отводится из почвы, опять же, растениями. Со временем вода возвращается в океан, чтобы продолжить круговорот. Виды образования от разных изменений Различают несколько видов круговоротов воды в природе: Большой, или мировой, круговорот — водяной пар, образовавшийся над поверхностью океанов, переносится ветрами на материки, выпадает там в виде атмосферных осадков и возвращается в океан в виде стока. В этом процессе изменяется качество воды: при испарении соленая морская вода превращается в пресную, а загрязненная — очищается. Малый, или океанический, круговорот — водяной пар, образовавшийся над поверхностью океана, сконденсируется и выпадает в виде осадков снова в океан. Внутриконтинентальный круговорот — вода, которая испарилась над поверхностью суши, опять выпадает на сушу в виде атмосферных осадков. В конце концов, осадки в процессе движения опять достигают Мирового океана. Скорость
Скорость переноса различных видов воды изменяется в широких пределах, так и периоды расходов, и периоды обновления воды также разные. Они изменяются от нескольких часов до нескольких десятков тысячелетий. Атмосферная влага, которая образуется при испарении воды из океанов, морей и суши, и существует в виде облаков, обновляется в среднем через восемь дней. Воды, входящих в состав живых организмов, восстанавливаются в течение нескольких часов. Это наиболее активная форма водообмена. Период обновления запасов воды в горных ледниках составляет около 1 600 лет, в ледниках полярных стран значительно больше — около 9 700 лет. ! Полное обновление вод Мирового океана происходит примерно раз в 2 700 лет. Термодинамика живых систем Состояние живых систем в любой момент времени (динамическое состояние) характерно тем, что элементы системы постоянно разрушаются и строятся заново. Этот процесс носит название биологического обновления. Для обновления элементов в живых системах требуется постоянный приток извне веществ и энергии, а также вывод во внешнюю среду тепла и продуктов распада. Это означает, что живые системы обязательно должны быть открытыми системами. Благодаря этому в них создается и поддерживается химическое и физическое неравновесие. Именно на этом неравновесии основана работоспособность живой системы, направленная на поддержание высокой упорядоченности своей структуры, а, значит, на сохранение жизни и осуществление различных жизненных функций. Кроме того, живая система, благодаря свойству открытости, достигает стационарности, т.е. постоянства своего неравновесного состояния. В изолированной системе (такая система не обменивается с внешней средой веществом и энергией), находящейся в неравновесном состоянии, происходят необратимые процессы, которые стремятся привести систему в равновесное состояние. Переход живой системы в такое состояние означает для нее смерть. Таким образом, открытость – одно из важнейших свойств живых систем. Весьма важным является вопрос о применимости законов термодинамики к живым системам. I закон (начало) термодинамики. Первый закон термодинамики гласит: изменение энергии системы (dE) равно количеству тепла (Q), полученному системой, плюс работа внешних сил (A), совершенная над системой dE = Q + A Для адиабатически изолированных систем (Q = 0, то есть обмена теплом с внешней средой не происходит) и замкнутых (А = 0, то есть внешние силы отсутствуют) dE = 0. Последнее утверждение является законом сохранения энергии: при всех изменениях, происходящих в адиабатически изолированных и замкнутых системах, полная энергия системы остается постоянной. Если рассматривать термодинамическую систему, состоящую только из живой системы, то закон сохранения энергии неприменим, так как живая система является открытой. Для термодинамической системы, включающей в себя живую систему и среду, с которой система обменивается энергией и веществом, закон сохранения энергии выполняется. Действительно, как показали опыты, общее количество энергии, которое получает организм за некоторый промежуток времени, вновь обнаруживается впоследствии в виде: а) выделяемого тепла; б) совершаемой внешней работой или выделяемых веществ; в) теплоты сгорания веществ, синтезированных за этот промежуток времени за счет энергии, поступившей извне. II закон (начало) термодинамики. Второй закон термодинамики утверждает, что в изолированной термодинамической системе энтропия никогда не может уменьшаться. Она равна нулю при обратимых процессах и может только увеличиваться при необратимых процессах. Здесь есть также определенная связь с упорядоченностью системы, а также с информацией (большая упорядоченность соответствует большему количеству информации). Можно говорить при этом о единстве природы информации и энтропии. Действительно, увеличение энтропии соответствует переходу системы из более упорядоченного в менее упорядоченное состояние. Такой переход сопровождается уменьшением информации, содержащейся в структуре системы. Беспорядок, неопределенность можно трактовать как недостаток информации. В свою очередь возрастание количества информации уменьшает неопределенность. Вспомним физический смысл энтропии. Все процессы, самопроизвольно протекающие в природе, необратимы и способствуют переходу системы в равновесное состояние, которое всегда характеризуется тем, что: а) в процессе этого перехода всегда безвозвратно выделяется некоторая энергия, и для совершения полезной работы она использована быть не может; б) в равновесном состоянии элементы системы характеризуются наименьшей упорядоченностью. Отсюда следует, что энтропия является как мерой рассеяния энергии, так и мерой неупорядоченности системы. Применение второго закона термодинамики к живым системам без учета того, что это открытые системы, приводит к противоречию. Действительно, энтропия должна всегда возрастать, то есть должна расти неупорядоченность живой системы. В то же время мы хорошо знаем, что все живые системы постоянно создают из беспорядка упорядоченность. В них создается и поддерживается физическое и химическое неравновесие, на котором основана работоспособность живых систем. В процессе развития каждого организма (онтогенеза), так же как и в процессе эволюционного развития (филогенеза), все время образуются новые структуры, и достигается состояние с более высокой упорядоченностью. А это означает, что энтропия (неупорядоченность) живой системы не должна возрастать. Таким образом, второй закон термодинамики, справед-ливый для изолированных систем, для живых систем, являющихся открытыми, неприменим. В течение времени жизни живой системы ее элементы постоянно подвергаются распаду. Энтропия этих процессов положительна (возникает неупорядоченность). Для ком<
|
||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2021-04-04; просмотров: 154; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.119.116.27 (0.013 с.) |