Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Задачи и методы генетики человекаСодержание книги
Поиск на нашем сайте
Задачи и методы генетики человека В частных разделах генетики исследуются особенности проявления общих закономерностей у разных видов организмов. Среди них ведущее положение занимает генетика человека, которая включает такие же разделы, как и общая генетика.
Методы генетики человека Гибридологический метод неприменим для изучения человека ввиду невозможности направленного скрещивания. Этот метод слабо применим и для многих крупных животных, поскольку потомство малочисленно, а время его развития сопоставимого с временем жизни экспериментатора.
Медицинская генетика Предметом медицинской генетики являются те направления генетики человека, которые изучают роль генетических факторов в этиологии и патогенезе заболеваний. Основной целью этих исследований является обеспечение здоровья не только настоящего, но и будущих поколений. Раздел медицинской генетики, используемый в клинической практике, называется клинической генетикой. Однако четкой границы между этими двумя направлениями не существует, и многие положения медицинской генетики, которые по началу рассматривались как чисто теоретические, через какое-то время находят применение в клинике. Интенсивно развивается фармакогенетика, т.е. изучение влияния генотипических особенностей пациентов на метаболизм лекарственных препаратов. С генетических позиций все болезни человека можно разделить на три класса:
В этиологии многофакторных заболеваний наряду с действием неблагоприятных внешних факторов существенно влияние не одного, а многих генов. Количество этих генов, формирующих наследственную предрасположенность, иногда исчисляется десятками или даже сотнями. К многофакторным заболеваниям относятся большинство наиболее распространенных болезней человека, таких как гипертония, атеросклероз, ишемическая болезнь сердца, бронхиальная астма, сахарный диабет, онкологические, аутоиммунные заболевания и др. Травмы и инфекции относятся к классу приобретенных заболеваний. Однако в последнем случае часто не удается полностью исключить влияния генетических факторов, определяющих дифференциальную чувствительность разных индивидуумов к действию инфекционных агентов. В этих случаях инфекционные заболевания рассматриваются как многофакторные. В задачи медицинской генетики входят:
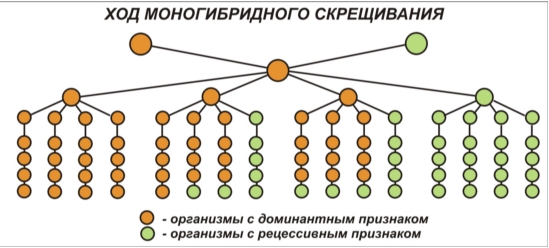
В настоящее время интенсивно изучают ассоциации разных генов человека с моногенными и многофакторными заболеваниями. Эти исследования являются основой для планомерной разработки совместно со специалистами разных медицинских профилей новых патогенетических и этиологических методов лечения наследственных заболеваний, а также предупреждения развития тех заболеваний, к которым у человека имеется генетическая склонность. Определение Генотип — это совокупность генов данного организма, а фенотип (от «фен» — признак) — это совокупность его признаков. Поскольку гены не всегда проявляются как признаки, организмы могут иметь одинаковый фенотип, но разные генотипы. Фенотип также зависит от взаимодействия генотипа и окружающей среды, то есть организмы с одинаковым генотипом могут иметь разные признаки (например, близнецы или растения при вегетативном размножении). 3. Мендель проводил точный количественный учет проявления признаков у потомства, разбивая его на группы по признакам и подсчитывая число особей (или семян) в каждой. Он оперировал в своей работе не качественными понятиями («больше — меньше»), а точными цифрами. Он анализировал эти цифры и старался усмотреть в них определенные математические соотношения. Это без преувеличения можно назвать первым синтезом математики и биологии, а в целом - переворотом в биологическом мышлении. 4. При анализе наследования Мендель всегда обращал внимание на каждый признак отдельно. Этот принцип и сегодня лежит в основе генетического анализа. Ранее исследователи пытались описать фенотип как целое, по всем признакам сразу. Это был тупиковый путь, так как в таком случае закономерности наследования становятся слишком сложными для того, чтобы их легко вычленить. Для описания наследования необходимо выделять отдельный признак и «не обращать внимания» на остальные. 5. Мендель брал в исходные скрещивания не любые растения, а только чистые линии. Определение Чистая линия — это совокупность организмов, которые на протяжении многих поколений скрещиваний друг с другом проявляют одинаковые признаки (не дают расщепления). Их получают путем близкородственных скрещиваний. Горох — самоопыляющееся растение, поэтому в данном случае чистые линии легко получаются путем самоопыления в течение нескольких поколений и отбора особей с постоянным проявлением признака в потомстве. Второй закон Менделя При скрещивании гибридов первого поколения между собой (самоопыления), которые были получены при искусственном перекрёстном опылении двух сортов гороха с альтернативными признаками, во втором поколении Мендель получил особи как с доминантными, так и с рецессивными состояниями признаков, т.е. наблюдал расщепление признаков. Проделав статистический анализ среди гибридов второго поколения, учёный обнаружил закономерность соотношений количеств потомков с разными признаками. Так, в опытах Менделя на 929 растений второго поколения оказалось 705 с пурпурными цветками и 224 с белыми. В опыте, в котором учитывался цвет семян, с 8023 семенами гороха, полученными во втором поколении, было 6022 жёлтых и 2001 зелёное, а с 7324 семенами, в отношении которых учитывалась форма семени, было получено 5474 гладких и 1850 морщинистых. Исходя из полученных результатов, Мендель пришел к выводу, что во втором поколении 75% особей имеют доминантное состояние признака, а 25% — рецессивное (расщепление 3:1). Эта закономерность получила название второго закона Менделя, или Закона расщепления.
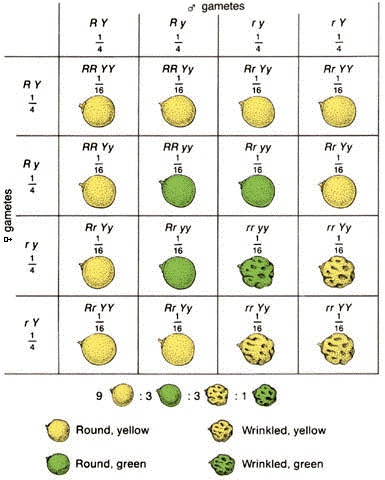
все рецессивные растения давали только рецессивное потомство, тогда как доминантные особи разделились на две группы: первая давала только доминантное потомство, а вторая и третья — доминантных и рецессивных потомков в отношении 3:1. Такие организмы несли два разных аллеля гена, так же как и их предок — гибрид первого поколения. Такие организмы называются гетерозиготными, в отличие от гомозиготных, несущих только один аллель данного гена. Таким образом, по своим наследственным свойствам (генотипу) гибриды первого поколения давали расщепление 1:2:1, а по внешним признакам (по фенотипу) 3:1. Мендель предположил также, что каждая гамета несёт только один аллель данного гена (принцип чистоты гамет). При скрещивании двух гетерозигот (Аа), в каждой из которых образуется два типа гамет (половина с доминантными аллелями А, половина с рецессивными а), необходимо ожидать четыре возможных сочетания. Яйцеклетка с аллелью А может быть оплодотворена с одинаковой долей вероятности как сперматозоидом с аллелью А, так и сперматозоидом с аллелью а; и яйцеклетка с аллелью а — сперматозоидом или с аллелью А, или с аллелью а. В результате получаются зиготы АА, Аа, Аа, аа или АА, 2Аа, аа. Третий закон Менделя Изучая расщепления при дигибридном скрещивании (скрещивании организмов, которые отличаются по двум признакам), Мендель обратил внимание на следующее обстоятельство. При скрещивании растений с жёлтыми гладкими (ААВВ) и зелёными морщинистыми (ааbb) семенами в первом поколении все горошины были гладкими и жёлтыми (AaBb), а во втором поколении появлялись новые комбинации признаков: жёлтые морщинистое (Ааbb) и зелёные гладкие (ааВb), которые не встречались в исходных формах. Соотношение форм по фенотипу было 9 жёлтых гладких (А-В-): 3 жёлтых морщинистых (А-bb): 3 зелёных гладких (аaВ-): 1 зелёный морщинистый (аabb). При этом соотношение жёлтых и зелёных оказалось 3:1, гладких и морщинистых тоже 3:1. Из этого наблюдения Мендель сделал вывод, что расщепление по каждому признаку происходит независимо от второго признака. В этом примере форма семян наследовалась независимо от их окраски. Эта закономерность получила название третьего закона Менделя, или закона независимого распределения генов.
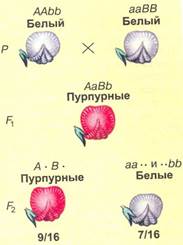
Третий Закон Менделя: Комплементарность К комплементарным, или дополняющим друг друга, генам относятся такие неаллельные гены, которые при совместном проявлении обусловливают развитие нового признака. На примере наследования окраски цветков у душистого горошка можно понять сущность комплементарного действия генов. При скрещивании двух растений с белыми цветками у гибридов F1 цветки оказались пурпурными. При самоопылении растений из F1 в F2 наблюдалось расщепление растений по окраске цветков в отношении, близком к 9:7. Пурпурные цветки были обнаружены у 9/16 растений, белые у 7/16.
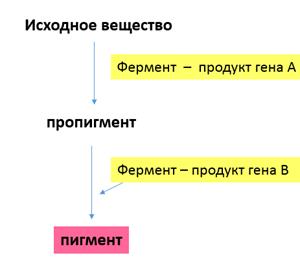
Объяснение такого результата состоит в том, что каждый из доминантных генов не может вызвать появление окраски, определяемой пигментом антоцианом. У душистого горошка есть ген А, обусловливающий синтез бесцветного предшественника пигмента — пропигмента. Ген В определяет синтез фермента, под действием которого из пропигмента образуется пигмент. Цветки душистого горошка с генотипом ааВВ и ААbb имеют белый цвет: в первом случае есть фермент, но нет пропигмента, во втором — есть пропигмент, но нет фермента, переводящего пропигмент в пигмент.
Проведем скрещивание двух растений душистого горошка с белыми цветками. В одной из линий, АAbb, есть доминантный аллель А, а у другой, ааBB, есть доминантный аллель В. У растений, имеющих доминантные гены А и В одновременно, есть и пропигмент (обеспечиваемый А), и фермент (обеспечиваемый В), необходимые для образования пурпурного пигмента. У гибридов F1 генотип AaBb, есть оба доминантных гена, поэтому они имеют пурпурные цветки. При самоопылении этих растений получаем F2. Результаты отражены в решётке Пеннета, розовым выделены генотипы тех растений, которые будут иметь пурпурные цветки.
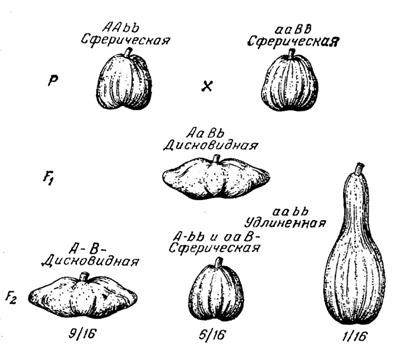
Комплементарное взаимодействие может давать не два, а три фенотипа. Это происходит в тех случаях, когда двойные рецессивные гомозиготы aabb по фенотипу отличаются от рецессивных гомозигот только по одному гену — aaBb, Aabb. В этом случае расщепление будет 9:6:1. Например, это наблюдается при наследовании формы плодов у тыквы:
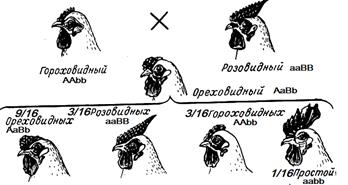
Возможно и возникновение четырёх фенотипов. Например, при скрещивании кур с различной формой гребня.
А_В_ — ореховидный А_bb — гороховидный aaB_ — розовидный aabb — простой (листовидный) Эпистаз Эпистаз — взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый — гипостатичным. Если эпистатичный ген не имеет собственного фенотипического проявления, то он называется ингибитором и обозначается буквой I. Эпистатическое взаимодействие неаллельных генов может быть доминантным и рецессивным. Доминантный эпистаз При доминантном эпистазе проявление гипостатичного гена (В, b) подавляется доминантным эпистатичным геном (I > В, b). Расщепление по фенотипу при доминантном эпистазе может происходить в соотношении 12:3:1, 13:3. Последний вариант возникает, когда рецессивная гомозигота по гипостатичному гену фенотипически неотличима от фенотипа I (например, обе формы неокрашены).
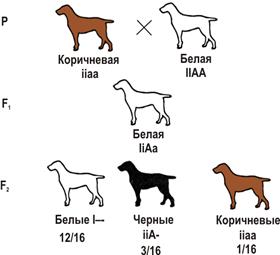
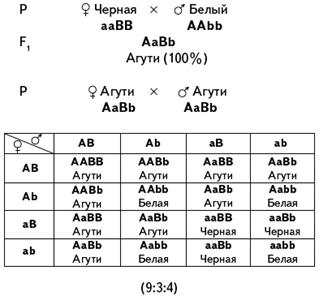
Наследование окраски шерсти у собак (пример доминантного эпистаза): A — черная окраска, а — коричневая, I — подавляет окраску, i — не подавляет. Рецессивный эпистаз Рецессивный эпистаз — это подавление рецессивным аллелем эпистатичного гена в гомозиготном состоянии аллелей гипостатичного гена (ii > В, b). Расщепление по фенотипу может идти в соотношении 9:3:4, 9:7. Последний вариант возникает, когда рецессивная гомозигота по гипостатичному гену фенотипически неотличима от фенотипа ii (например, обе формы неокрашены). Рецессивный эпистаз проявляется при наследовании окраски шерсти у домовых мышей. А — окраска агути (рыжевато-серая) а — черная окраска В — способствует проявлению окраски b — супрессор (подавляет действие А и а) Мыши с генотипом А-bb и ааbb имеют одинаковый фенотип — все белые.
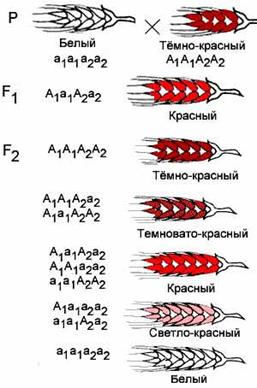
Полимерия Полимерия — взаимодействие неаллельных генов, при котором степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного гена имеют одинаковый нижний индекс. Например, у пшеницы А — тёмно-красный цвет зёрен, а — белый цвет зёрен. За цвет отвечают два гена — 1 и 2. Первый ген может быть представлен аллелями А1 и а1, второй — аллелями А2 и а2. В зависимости от того, каких аллелей больше — А или а, оттенок зёрен будет изменяться.
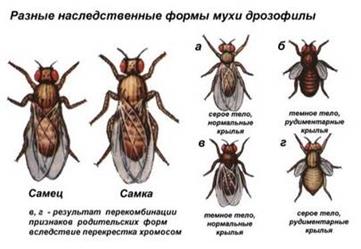
Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным. При кумулятивной (накопительной) полимерии степень проявления признака зависит от числа доминантных аллелей всех генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак. При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление по фенотипу происходит в соотношении 15:1 для двух генов, 63:1 для трёх генов и т.д. Например, цвет кожи у людей зависит от четырёх генов, поэтому от брака двух мулатов практически никогда не рождаются дети с белой кожей. Разберём решение задач с неаллельным взаимодействием генов. Задача 1 При скрещивании двух божьих коровок с красными надкрыльями все гибриды первого поколения имели оранжевые надкрылья. Их скрестили между собой и получили во втором поколении 115 жуков с оранжевыми надкрыльями, 79 с красными надкрыльями и 13 с жёлтыми надкрыльями. Как наследуется цвет надкрылий? Решение Так как в первом поколении все потомки одинаковы, родители гомозиготны. Появление во втором поколении третьей окраски и численные соотношения указывают на то, что это дигибридное скрещивание. Значит, в потомстве имеется 16 генотипических классов. Общее число потомков второго поколения = 115 + 79 + 13 = 207. Размер генотипического класса = 207: 16 = 13. С оранжевой окраской 115: 13 = 9 генотипических классов. С красной окраской 79: 13 = 6 генотипических классов. С жёлтой окраской 13: 13 = 1 генотипический класс. Таким образом, во втором поколении наблюдаем расщепление 9:6:1, что соответствует комплементарному взаимодействию генов. Родительские формы ААbb и ааВВ, гибриды первого поколения АаВb. Во втором поколении 9 А__В_: 6 (ааВ_ + А_bb):1 ааbb. Задача 2 При скрещивании собак чистых линий с коричневой и белой окраской все потомки были белыми. Во втором поколении было получено 167 белых щенков, 43 чёрных и 14 коричневых. Как наследуется окраска? Решение Наличие трёх фенотипов и численные отношения, далёкие 1:2:1 указывают на дигибридное скрещивание. Определяем размер генотипического класса. (167 + 43 + 14): 16 = 224: 16 = 14. Белые — 167: 14 = 12, чёрные 43: 14 = 3, коричневые 14: 14 = 1. Соотношение фенотипов 12:3:1, что соответствует доминантному эпистазу. Генотипы родителей aaii (коричневый) и AAII (белый), первое поколение AaIi. Во втором поколении 12__I_(белые) +3 A_ii (чёрные) + 1 aaii (коричневые). Опыты Моргана Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, — над геном недоразвитых).
При проведении анализирующего скрещивания самки F1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% — серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% — черные длиннокрылые и 8,5% — серые с зачаточными крыльями). А — серое тело а — черное тело В — нормальные крылья b — недоразвитые крылья
Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, соединены между собой. Оказалось, что гены образуют группы сцепления, т.е. гены одной группы наследуются сцеплено, а гены разных групп — независимо. Все гены одной хромосомы образуют группу сцепления и наследуются совместно. Количество групп сцепления соответствует гаплоидному набору хромосом. Поэтому Морган предположил, что гены локализованы на хромосомах. Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Группы сцепления разрушаются при кроссинговере, когда происходит обмен участками гомологичных хромосом в профазу I мейоза. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними. Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АВ и аb, а отцовский — один тип — аb. Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип АаВb и ааbb. Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb. Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток — мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Аb и аВ, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1. Гибридные особи, имеющие иное сочетание признаков, чем у родителей называются рекомбинантами. Расстояние между генами измеряется в морганидах — условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам. Задача У кукурузы гладкая поверхность зёрен доминирует над морщинистой,неокрашенные зёрна рецессивный признак. Скрестили линию кукурузы с гладкими окрашенными зёрнами и с морщинистыми неокрашенными. В первом поколении все растения имели гладкие окрашенные зёрна. Гибридов первого поколения скрестили с родительским сортом с неокрашенными морщинистыми зёрнами. Получили 4152 растения с гладкими окрашенными зёрнами, 149 с морщинистыми окрашенными, 152 с гладкими неокрашенными и 4166 с морщинистыми неокрашенными. Как наследуются признаки.
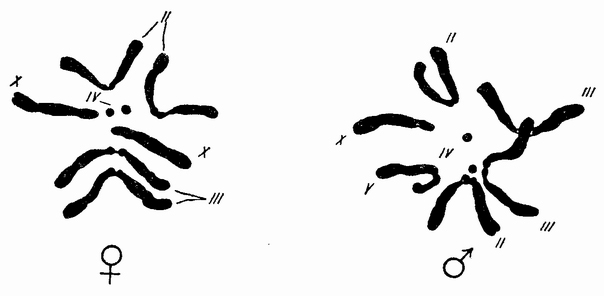
Подавляющее большинство видов животных представлено особями двух полов — мужского и женского. Расщепление по половой принадлежности происходит в соотношении 1:1. Иными словами, у всех видов численность самцов и самок приблизительно одинакова. Еще Г. Мендель обратил внимание на то, что такое расщепление в потомстве по какому-либо признаку наблюдается в тех случаях, когда одна из родительских особей была гетерозиготой (Аа) по этому признаку, а вторая — рецессивной гомозиготой (аа). Было сделано предположение, что один из полов (тогда было неясно, какой именно) гетерозиготен, а второй гомозиготен по гену, который определяет пол организма. Современная теория наследования пола была разработана Т. Морганом и его сотрудниками в начале XX в. Им удалось установить, что самцы и самки различаются по набору хромосом. У мужских и женских организмов все пары хромосом, кроме одной, одинаковы и называются аутосомами, а одна пара хромосом, называемых половыми, — у самцов и самок различается. Например, и у самцов, и у самок дрозофил в каждой клетке по три пары аутосом, а вот половые хромосомы различаются: у самок — по две Х-хромосомы, а у самцов X и Y (рис 1). Пол будущей особи определяется во время оплодотворения. Если сперматозоид содержит Х-хромосому, то из оплодотворенной яйцеклетки разовьется самка (XX), а если в сперматозоиде содержалась половая Y-хромосома — то самец (ХY). Соотношение полов при таком скрещивании теоретически всегда будет 1:1. Так как у самок дрозофил образуются только яйцеклетки, содержащие половые Х-хромосомы, то женский пол у дрозофил называют гомогаметным. У самцов дрозофил образуются в равном соотношении сперматозоиды либо с Х-, либо Y-половыми хромосомами. Поэтому мужской пол у дрозофил называется гетерогаметным. У многих видов живых существ, например у ракообразных, земноводных, рыб, большинства млекопитающих (в том числе и человека), женский пол гомогаметный (XX), а мужской — гетерогаметный (ХY). У людей Y-хромосома, определяющая мужской пол, передается от отца к сыну в момент оплодотворения. Таким образом, пол младенца зависит только от того, какая из половых хромосом попала в зиготу от отца. В У-хромосоме человека находятся гены белков, необходимых для нормального развития мужских половых желез. Эти железы очень быстро начинают выделять мужские половые гормоны, определяющие формирование всей половой системы мужчины. Если же в оплодотворении участвовал сперматозоид с Х-хромосомой, то в клетках развивающегося зародыша Y-хромосома отсутствует, значит, нет и кодируемых ей «мужских» белков. Поэтому в зародыше девочки развиваются яичники и женские половые пути. Итак, у дрозофилы и человека женский пол является гомогаметным, и общая схема наследования пола у двух этих видов одинакова. У некоторых видов живых существ хромосомное определение пола совсем другое. Например, у птиц и рептилий — гомогаметны самцы (ZZ), а самки — гетерогаметны (ZW). У некоторых насекомых (например у сверчков) у самцов в хромосомном наборе лишь одна половая хромосома (ХО), а самки — гомогаметны (XX). У пчел и муравьев половых хромосом нет, и самки имеют в клетках тела диплоидный набор хромосом, а самцы, развивающиеся партеногенетически (из неоплодотворенных яйцеклеток), — гаплоидный набор хромосом. Естественно, что в этом случае развитие сперматозоидов у самцов идет без мейоза, так как уменьшить число хромосом менее гаплоидного набора невозможно. У крокодилов половые хромосомы не обнаружены. Пол зародыша, развивающегося в яйце, зависит от температуры окружающей среды: при высоких температурах развивается больше самок, а в том случае, если прохладно, — больше самцов.
Основные закономерности наследования признаков были впервые сформулированы в работах австрийского исследователя Грегора Менделя. Менделя считают отцом генетики — науки о закономерностях наследственности и изменчивости организмов. Он использовал в своей работе гибридологический метод генетики. Этот метод заключается в скрещивании организмов с определенными признаками и анализе проявления признаков у потомства. Предшественники Менделя уже пытались установить основные закономерности наследственности. Они занимались скрещиванием растений, реже животных и наблюдали такие явления, как доминирование и расщепление, однако не смогли сделать достаточно общих выводов из наблюдений. Что же позволило работе Менделя стать революцией в биологии, и каковы ее основные принципы? 1. Одной из важнейших составляющих успеха Менделя было то, что он скрещивал сорта гороха, которые различались парами альтернативных признаков. Альтернативные признаки имеют четко различимые взаимоисключающие проявления без промежуточных форм по принципу «или — или». Например:
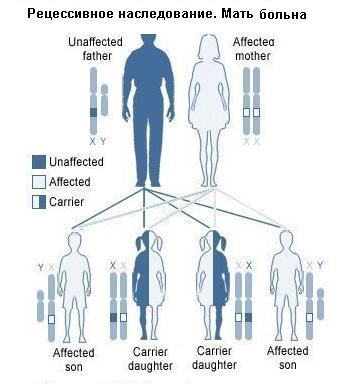
2. Второй составляющей работы Менделя является анализ генотипа и фенотипа организмов. Определение Генотип — это совокупность генов данного организма, а фенотип (от «фен» — признак) — это совокупность его признаков. Поскольку гены не всегда проявляются как признаки, организмы могут иметь одинаковый фенотип, но разные генотипы. Фенотип также зависит от взаимодействия генотипа и окружающей среды, то есть организмы с одинаковым генотипом могут иметь разные признаки (например, близнецы или растения при вегетативном размножении). 3. Мендель проводил точный количественный учет проявления признаков у потомства, разбивая его на группы по признакам и подсчитывая число особей (или семян) в каждой. Он оперировал в своей работе не качественными понятиями («больше — меньше»), а точными цифрами. Он анализировал эти цифры и старался усмотреть в них определенные математические соотношения. Это без преувеличения можно назвать первым синтезом математики и биологии, а в целом - переворотом в биологическом мышлении. 4. При анализе наследования Мендель всегда обращал внимание на каждый признак отдельно. Этот принцип и сегодня лежит в основе генетического анализа. Ранее исследователи пытались описать фенотип как целое, по всем признакам сразу. Это был тупиковый путь, так как в таком случае закономерности наследования становятся слишком сложными для того, чтобы их легко вычленить. Для описания наследования необходимо выделять отдельный признак и «не обращать внимания» на остальные. 5. Мендель брал в исходные скрещивания не любые растения, а только чистые линии. Определение Чистая линия — это совокупность организмов, которые на протяжении многих поколений скрещиваний друг с другом проявляют одинаковые признаки (не дают расщепления). Их получают путем близкородственных скрещиваний. Горох — самоопыляющееся растение, поэтому в данном случае чистые линии легко получаются путем самоопыления в течение нескольких поколений и отбора особей с постоянным проявлением признака в потомстве. Наследование дальтонизма Известны нарушения цветового зрения, так называемая цветовая слепота. В основе появления этих дефектов зрения лежит действие ряда генов. Красно-зеленая слепота обычно называется дальтонизмом. Еще задолго до появления генетики в конце XVIII и в XIX в. было установлено, что цветовая слепота наследуется согласно вполне закономерным правилам. Так, если женщина, страдающая цветовой слепотой, выходит замуж за мужчину с нормальным зрением, то у их детей наблюдается очень своеобразная картина перекрестного наследования. Все дочери от такого брака получат признак отца, т. е. они имеют нормальное зрение, а все сыновья, получая признак матери, страдают цветовой слепотой. В том же случае, когда наоборот, отец является дальтоником, а мать имеет нормальное зрение, все дети оказываются нормальными. В отдельных браках, где мать и отец обладают нормальным зрением, половина сыновей может оказаться пораженными цветовой слепотой. В основном наличие цветовой слепоты чаще встречается у мужчин. Э.Вильсон объяснил наследование этого признака, предположив, что он локализовал в Х-хромосоме и что у человека гетерогаметным (XY) является мужской пол. Становится вполне понятным, что в браке гомозиготной нормальной женщины (ХD ХD) с мужчиной дальтоником (ХDY) все дети рождаются нормальными. Однако при этом, все дочери становятся скрытыми носителями дальтонизма, что может проявиться в последующих поколениях.
Наследование дальтонизма Известны нарушения цветового зрения, так называемая цветовая слепота. В основе появления этих дефектов зрения лежит действие ряда генов. Красно-зеленая слепота обычно называется дальтонизмом. Еще задолго до появления генетики в конце XVIII и в XIX в. было установлено, что цветовая слепота наследуется согласно вполне закономерным правилам. Так, если женщина, страдающая цветовой слепотой, выходит замуж за мужчину с нормальным зрением, то у их детей наблюдается очень своеобразная картина перекрестного наследования. Все дочери от такого брака получат признак отца, т. е. они имеют нормальное зрение, а все сыновья, получая признак матери, страдают цветовой слепотой. В том же случае, когда наоборот, отец является даль
|
|||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2021-02-07; просмотров: 203; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.12.233 (0.019 с.) |