Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Особенности зрения у собак некоторых пород с крайними морфологическими характеристиками (по Е. П. Копенкину)Содержание книги
Поиск на нашем сайте
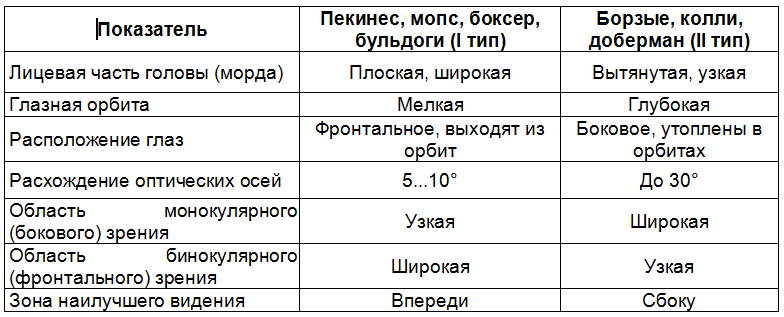
Разумеется, животные других пород с промежуточными морфологическими характеристиками обладают средним зрением. Все это необходимо учитывать при требовании от собак выполнения каких‑то действий, связанных с их зрительными способностями. Например, животные I типа превосходят животных II типа по остроте зрения и точности определения дистанции близко перед собой, что связано с более развитой конвергенцией оптических осей, но уступают им по панорамности обзора и улавливанию подвижности объекта на большом расстоянии. Еще одним показателем зрения является перспектива, которая обусловлена расстоянием от земли до глаз, что зависит от роста собаки. Разумеется, у низкорослых животных такая перспектива будет значительно меньше, чем у высокорослых. Можно сравнить крайние варианты пород по росту: брабансон, брюссельский и бельгийский гриффоны – 18…20 см и немецкий дог – до 80 см, ирландский волкодав – 86 см [82]. Более сложный вариант проявления зрения у собак – это способность к дифференцировке предметов, например геометрических фигур. Специальными исследованиями выявили достаточно высокое развитие у них этой способности. Так, в работе ученика И. П. Павлова М. М. Губергрица (1917) собаки отличали круг от эллипса с соотношением полуосей 8:9 [85]. В работе W. H. Karn et al. (1932) собаки быстро научились различать горизонтальные и вертикальные линии и более медленно – поставленный на основание и перевернутый на вершину треугольники. При этом не имели значения размеры объектов и были они контурными или закрашенными.
Интересную работу провели в одном из московских клубов служебного собаководства. Собаку научили садиться при виде круга, ложиться при показе треугольника вершиной вниз и стоять при показе стоящего на основании квадрата. Все фигуры были закрашены. Правильное выполнение визуальных команд началось после 2…3 предъявлений. И все‑таки не обходилось без ошибок: даже после научения правильное выполнение составило в целом 87 %. Затем положение фигур стали менять, но с прежним выполнением визуальных команд: сидеть, лежать и стоять. Повернули квадрат, поставив его на вершину, а треугольники повернули вершинами вниз, влево и вправо. Повернутые треугольники собака узнала сразу, а перевернутый квадрат – со второго предъявления. Затем те же опыты проводили с большими фигурами, а также предъявляли контурные круги и треугольники разного размера (ранее они были закрашены). Во всех вариантах правильное выполнение составило 100 %, в том числе при различении фигур по размерам. Потом показывали те же фигуры, но заштрихованные. Круг и треугольник собака узнала сразу, а квадрат – со второго предъявления. Далее провели опыт, являющийся смешением всех предыдущих: те же фигуры были разных размеров, положений и качества поверхности. В ответ на такую смесь вариантов число правильных ответов снизилось до 93 %. В следующей группе опытов изменили поведенческую реакцию собаки: научили подавать лапу при показе заштрихованного круга. При показе заштрихованного и не заштрихованного кругов она начала их правильно различать со второго предъявления. В предыдущем аналогичном опыте та же собака начала различать такие же круги, но с посадкой, сразу. Далее опыт усложнили: на показ круга с вертикальной штриховкой собака должна была подавать правую лапу, на круг с горизонтальной штриховкой – левую. В результате она очень часто ошибалась; даже после научения правильное выполнение составило лишь 57 %. Отсюда можно сделать вывод: собака не различала наклоны штриховки, хотя в предыдущем опыте она успешно различала заштрихованную и незаштрихованную фигуры. Таким образом, собаки не только различают геометрические фигуры, даже очень похожие, но и узнают эти фигуры при изменении их характеристик, т. е. собаки способны отвлекаться от частностей, удерживая во внимании сущность. А это, как известно, составляет основу элементарной рассудочной деятельности, является проявлением мышления. Однако у такой способности оказался предел: отличение треугольников вершиной вверх и вниз; отличение квадратов в разных положениях; и совсем непреодолимое препятствие – различение одной и той же фигуры с разнонаправленной штриховкой. Кроме того, на успех решения задачи влияет способ ответного (сигнального) действия собаки, а также побочные характеристики различаемых объектов.
Все проявления зрения у собак предназначены для восприятия отдельных, хотя и взаимосвязанных, проявлений окружающего мира. Но все они обусловливаются обобщающей и самой сложной функцией зрения – его остротой, т. е. способностью ясно видеть мелкие объекты или их детали. Острота зрения – наименьшее расстояние между предметами, при котором они воспринимаются раздельно [20], или наименьший угол, под которым две точки видны раздельно [47, 50], а также наибольшая частота вспышек, при которой они не сливаются в сплошной свет. По данным электроретинограммы (ЭРГ), у анестезированных собак палочки способны раздельно улавливать вспышки с максимальной частотой 20 Гц. При более сильной освещенности, когда активизируются колбочки, слияние вспышек происходило при частоте 70 Гц. Например, 4 бигля, приученные нажимать педаль при вспышках, раздельно воспринимали их при более высокой максимальной частоте (до 80 Гц) и меньшей освещенности, чем при определении путем снятия ЭРГ [108]. У людей раздельное восприятие палочками максимальной частоты вспышек (70 Гц) примерно такое же, как у собак, а восприятие колбочками немного ниже (50…60 Гц). Остроту зрения измеряют и другими методами. Один из них – метод Снеллена: он заключается в сравнении стандартного (постоянного) расстояния, с которого испытуемый ясно видит самые мелкие элементы, с расстоянием, с которого субъект с нормальным зрением, принимаемый за эталон, ясно видит то же. Его наиболее часто применяют в опытах с собаками, в частности, используют диаграмму Снеллена, в которой на экране меняют расстояние между темными и светлыми полосами до тех пор, пока испытуемый продолжает их видеть раздельно. В поведенческих опытах острота зрения у собак при сильной освещенности (37 лк) составляла в среднем 0,21. При определении у 3 биглей электропотенциала в коре головного мозга в результате освещения сетчатки острота зрения оказалась равной 0,235…0,308, а по ЭРГ – 0,40…0,43. Потом опыт усложнили. Горизонтальные полосы двигались с одновременным изменением расстояния между ними. Определяли минимальное расстояние, когда начинался нистагм (непроизвольное, ритмичное, пилообразное движение глаз в ответ на зрительные или вестибулярные раздражения). Острота зрения составила 0,2. Далее ЭРГ сравнивали по площадям сетчатки: 15° в центре и 15° тороидально вокруг него. Острота зрения в этих участках составила 0,143 и 0,085 соответственно (R. Orfi et al, 1993). Все эти исследования подтвердили низкую остроту зрения у собак (в среднем 0,267), так как при наличии у них панорамного зрения световоспринимающая способность сетчатки снижается с удалением от центра в связи с уменьшением количества фоторецепторов на периферии. Японские исследователи определяли остроту зрения у трех собак породы шиба в поведенческих опытах. Их приучили к подкормке при виде обычного круга. Потом показывали круг Ландолта, т. е. с разрывом в каком‑то месте окружности (в данном случае внизу), на который собаки не должны реагировать так, как на нормальный круг. При этом меняли расстояние до кругов, их диаметр, ширину окружности и разрыва в ней. За максимальную остроту зрения принимали ситуацию, когда круги правильно различались в 3 сеансах подряд из 21 на уровне не менее 70 % предъявлений (р<0,05). Острота зрения у собак оказалась равной 0,23; 0,24 и 0,33 [125]. Еще более естественные поведенческие опыты описывают P. E. Miller, С. J. Murphy (1995). Они определяли чувствительность сетчатки по угрожающей реакции собаки на движение руки поперек ее поля зрения или способности следовать за движущимся хлопковым мячиком. Так вот, в самых трудных экспериментальных условиях, когда острота зрения падала ниже 0,05, у собак еще сохранялись положительные зрительные реакции. Человека с таким зрением считают слепым. Отсюда следует обоснованный вывод о том, что различение мелких деталей для жизнедеятельности собак (даже рабочих) менее важно, чем для большинства людей. Сравнивали значение для реальной жизни собак способностей различать мелкие детали при ярком свете и видеть в сумерках. Низкая острота зрения у собак предопределена морфофизиологическими характеристиками глаз: малое число колбочек даже в месте наилучшего видения и их низкая светочувствительность и отсутствие связи каждой из них с центром (G. Schubert, 1950); большое число палочек с их значительной пространственной суммацией и потому низкой разрешающей способностью; слабая конвергенция оптических осей. И все это в сочетании со слабостью такого мощного корректирующего фактора, как аккомодация. Все вместе взятое усугубляется расплывчатостью отраженного тапетумом света внутри глаза. Зрительные представления о любом предмете значительно расширяются и обогащаются, если помимо описанных выше его характеристик воспринимается цвет. Последнее возможно лишь через способность различать цвета, для проявления которой необходимо цветовое зрение (ЦЗ). Из всех чувств собаки наличие его остается наиболее спорным, хотя изучается более 100 лет. Это является следствием того, что несмотря на обычность и однотипность методов исследования ЦЗ (выработка дифференцировочного УР на подкрепляемые и неподкрепляемые цвета), получали неоднозначные результаты. Да и объясняли их не всегда адекватно и объективно. Поэтому до сих пор и нет единого мнения о наличии (или отсутствии) ЦЗ у собак. Одной из причин служит трудность проведения опыта в «чистом» виде. Изучаемые цветные объекты могут обладать совокупностью неуловимых для экспериментатора (но уловимых собакой) признаков: размеры и форма, подвижность, качество поверхности, а если используется свет, то яркость, температура, мигание и т. д., не говоря уже об окружающей обстановке и индивидуальных особенностях животных. Да и сам цвет может иметь разные насыщенность, оттенки. Поэтому выводы по результатам опыта иногда были ошибочными. Вот тому два примера, начиная с первого исследования, которое провел F. Graber (1884). Оно заключалось в том, что девять собак разных пород (половина – фокстерьеры) имели выбор свободного входа в светло‑красную и темно‑синюю камеры. Лишь две собаки (фокстерьер и пудель) чаще заходили в темно‑синюю, чем в светло‑красную. Остальные заходили в любую камеру, но когда яркость окраски камер выровняли, то «положительные» собаки одинаково часто заходили как в синюю, так и в красную камеру. Тем не менее собаки по‑разному реагируют на объекты различного цвета, хотя совершенно очевидно, что это больше связано с интенсивностью окраски [74]. Еще раньше такое мнение высказали W. Nagel, F. Himstedt (1902): в большинстве случаев, где исследователи находят различение цветов, различная реакция животных может быть приписана именно разнице в степени освещенности, а не разнице в цвете [74]. Другой пример. Некоторые исследователи зажигали перед мордой собаки фонари с красным и синим стеклами. Причем в первом случае давалась подкормка, во втором не давалась. В результате легко вырабатывалась дифференцировка двух цветов. Однако при этом не учли, что красное стекло нагревается сильнее, чем синее, а разницу в температуре собака могла ощущать. Тогда перед фонарями поместили водяной светофильтр, поглощающий исходящую от них теплоту. В результате легко выработавшаяся дифференцировка цветов исчезла [76]. Следовательно, собака ориентировалась не на цвет стекла, а на его температуру. В других опытах подобные ошибки старались не допускать или, по крайней мере, учитывать, но тем не менее результаты исследований отличаются неоднозначностью. Поэтому вопрос о цветовом зрении собак до сих пор остается открытым. Способность собак различать цвета изучали на основе слюноотделительных и поведенческих рефлексов. В качестве раздражителей применяли свет разных цветов и разноцветные предметы. Так, собаку с фистулой слюнной железы подкармливали одновременно с показом на экране красного квадрата от находящегося сзади фонаря [73, 74]. После 65…70 таких совпадений красный цвет без подкормки становился возбудителем слюноотделения. Затем на экране поочередно показывали квадраты зеленого, синего, фиолетового и белого цветов, но без подкормки. Тем не менее во всех этих случаях выделялось такое же количество слюны, как при показе красного квадрата. Если же после многократного показа фиолетового и синего или фиолетового, синего и зеленого квадратов показывали красный, который раньше совпадал с подкормкой, он тоже не вызывал слюноотделения. Однако при восстановлении возбуждающего действия красного квадрата показанный за ним зеленый квадрат опять стал вызывать слюноотделение. В этих опытах проявились иррадиации возбуждения и торможения (см. гл. 1). С уменьшением интенсивности каждого цвета путем увеличения расстояния экрана от собаки с 1 до 3 м, а также при показе черного квадрата после квадратов любого цвета повышалось количество отделяемой слюны. Следовательно, на слюноотделение действовал не столько цвет, сколько его интенсивность. Недостоверные результаты были получены G. Nicolai (1907) при изучении на основе слюноотделения способности различать цвета у двух собак. В ответ на показ красной или зеленой чашки при различных степенях освещенности они очень скоро совершенно запутались, поэтому не удалось с несомненностью установить способность различения цветов собаками. На основании результатов обеих работ Л. А. Орбели считал, что собака не может реагировать на разные цвета различно, так как во всех случаях ее реакция определялась всецело изменениями в интенсивности освещенности. В другом опыте Л. А. Орбели предлагал двум собакам с фистулой слюнной железы геометрические фигуры, различающиеся по направлению движения, форме, внешнему очертанию, качеству поверхности, сочетая или не сочетая их с подкормкой. Во всех случаях четко проявлялась дифференцировочная слюноотделительная реакция. Обобщая результаты этой и предыдущей работы по различению цветов[2], автор заключил, что ориентирование собаки в области световых явлений определяется тремя факторами (колебаниями интенсивности света, даже очень слабыми, движением и формой предметов) и что собака лишена такого могучего ориентировочного средства, как цветовая гамма. Потребовалось много времени, чтобы Л. А. Орбели (1938), став известным физиологом, академиком, пришел к выводу, что нельзя говорить, что собака не различает цветов, на том основании, что не обнаруживается разница в слюнном пищевом рефлексе[3]: «… разница между двумя моментами: способностью анализаторов более или менее тонко дробить внешние явления и способностью животного связывать отдельные детали с эффекторной деятельностью и проявлять различную деятельность под влиянием различных раздражителей. Можно хорошо различать раздражения, но не уметь различно на них реагировать». Таким образом, было подтверждено наличие цветового зрения у собак, что вполне подтверждается преобладанием положительных результатов по его изучению. Однако приблизительно в этот же период были опубликованы результаты исследований по выработке у собак поведенческих условных рефлексов на основе цвета: частично результаты были отрицательными, но в большинстве случаев – положительными. Как пример можно привести две работы, где в качестве раздражителя тоже использовался свет разного цвета, но подкрепления были разные. В первой работе на переднюю лапу собаки наносили электроболевое раздражение при красном свете и не наносили при свете других цветов. После многократных сочетаний она отдергивала лапу только при зажигании красного света [13]. Во второй работе двух собак научили подходить к кормушке при красном свете различной яркости и не подходить при зеленом и синем свете. Было сделано заключение, что у собак несомненно существует способность различать цвета, но в отношении быстроты развития дифференцировки встречаются значительные индивидуальные различия. В следующем опыте Л. А. Орбели (1913) позволял собаке подходить к корму при свете красного фонаря и не позволял подходить при свете сначала сине‑фиолетового, потом зеленого фонаря. Различение красного и сине‑фиолетового цветов вырабатывалось очень медленно и трудно, а различение красного и зеленого цветов вырабатывалось еще труднее. Но даже сформировавшееся различение было очень нестойким: нескольких дней перерыва в работе оказалось достаточно, чтобы оно исчезло. Правда, потом различение можно было восстановить и сделать это значительно легче, чем выработать вначале. Интенсивность любого цвета играла определяющую роль в скорости и правильности его выбора при решении о возможности или запрете подхода к кормушке. Исходя из результатов своих работ и противоречивых результатов других исследователей, автор пришел к заключению, что у тех собак, у которых обнаруживалось различение цветов, оно является рудиментарным, поскольку вырабатывается с большим трудом, требует значительной силы света и отличается нестойкостью. В других исследованиях использовали разноцветные предметы, но применяли их по‑разному. Первая серия опытов заключалась в предоставлении собакам возможности заходить в камеру или подходить к объекту, обозначаемому цветом, который сочетался с подкормкой, и не заходить в камеру и не подходить к объекту, обозначаемому цветом, который не сочетался с подкормкой. Во всех случаях расположение цветоносителей постоянно меняли (Е. М. Smith, 1912). Семь собак научили заходить в камеру с красной картонкой. Затем им предоставлялся свободный выбор входа в четыре камеры: на верху одной из них была прикреплена известная картонка красного цвета, на следующих двух – картонки других цветов или серые, а четвертая камера была без картонки. В конце концов животные в большинстве случаев входили в камеру с красной картонкой. Однако такое предпочтение проявилось только у трех собак (фокстерьер, вельштерьер и уиппет) и не проявилось у четырех остальных (шипперке, ирландский терьер и два фокстерьера). Причем это предпочтение легко затормаживалось при изменениях яркости и расположения цветных картонок. Надо отдать должное исследователю: и до, и после мало кто проводил подобные опыты на таком количестве собак. Был сделан вывод, что различение цветов собакой чрезвычайно нестойкое. Еще раньше А. Самойлов и А. Феофилактова (1907) указывали, что собака пользовалась способностью различать цвета (зеленый и серый) после обучения лишь при хорошо знакомых условиях. Если же поставленная ей задача чем‑нибудь усложнялась, то она руководствовалась в своем поведении не цветом, а другими признаками предметов. Аналогичный опыт провела В. К. Шепелева (1954), только «положительным» цветом (сигналом для захода в камеру) был зеленый, а «отрицательным» – красный, фиолетовый, синий и оранжевый. Для достижения правильных заходов (100 %) двум беспородным собакам потребовались 650…700 сочетаний зеленого диска с дисками других цветов. Такая наученность нарушилась заменой зеленого диска темно‑зеленым и еще больше – светло‑зеленым. Однако путем длительной тренировки восстанавливалась правильность заходов в камеры с новыми дисками. Во всех ситуациях длительность обучения у животных различалась. По такому же принципу А. А. Бабушкин (2002) научил 10 собак разных пород подходить к чашке с зеленой карточкой (с одновременной командой «Зеленый!») и не подходить к чашкам с красной и синей карточками. Только при этом условии они получали подкормку и только из чашки с зеленой карточкой. Но даже через 8 нед правильность подходов составляла 30…90 %. При замене зеленой карточки светло‑зеленой даже у собак с наибольшим исходным числом правильных подходов (77…90 %) увеличилось число ошибок. Путем длительной тренировки так и не удалось восстановить исходное число правильных подходов: в лучшем случае у трех собак из пяти оно достигло 76…87 %. Подобное исследование провели на двух собаках породы шиба. Они научились подходить к кормушке с красной, синей или желтой карточкой (в этих случаях они получали подкормку) и не подходить к кормушке с серой карточкой [125]. Индивидуальные особенности собак здесь проявились не только в длительности тренировки для совершения правильных действий, но и в разной затрате времени для отличения каждого из трех цветов от серого. Например, для одной из них для отличения красного, зеленого и синего цветов от серого потребовались соответственно 3, 6 и 9 испытаний. Вторая серия опытов заключалась в том, что собака должна выбрать из массы разноцветных предметов такой, который соответствует цвету, показанному экспериментатором. Впервые такие работы были проведены в начале XX в. W. Nagel, F. Himstedt (1902) научили пуделя брать с пола синюю, оранжевую или желтую карточку в ответ на показ карточек аналогичных цветов. Другой пудель научился выбирать красный шар в массе зеленых и синих, затем – шар любого цвета в соответствии с показанным экспериментатором. Но самое интересное заключалось в том, что необходимый цвет потом ему называли. Несмотря на длительность обучения (10 нед), был сделан вывод: у собаки можно выработать реакцию именно на определенный цвет, независимо от освещенности. К такому же выводу пришли А. Самойлов и A. Феофилактова (1907): продолжительными упражнениями можно заставить собаку различать цвета. В их эксперименте собака отличала зеленый цвет от серого разных степеней освещения. У В. Л. Дурова (1924) во время сеансов экспериментальной дрессировки немецкая овчарка приносила разнообразные предметы разных цветов и разной яркости в соответствии с показанным экспериментатором. Трех спаниелей [120] научили выбирать красную пластиковую тарелку, из которой производилась подкормка, среди комплекта серых, из которых ее не подкармливали. Потом подкормка давалась из рук человека за выбор красной тарелки среди серых. После выработки прочного условного рефлекса серые тарелки поочередно заменяли голубыми, зелеными и желтыми. Позже они, тоже поочередно, служили «положительными» объектами, т. е. за их выбор среди комплектов других цветов собаки получали награду (из рук). Причем тарелки всех цветов имели разную яркость, но во время одного сеанса яркость тарелок всех цветов была одинаковой. В итоге все собаки безошибочно различали цвета любой яркости. Но выявились индивидуальные особенности: одна из них с трудом выбирала красный цвет в зеленом комплекте, другая – зеленый цвет в желтом комплекте. И наконец, третья серия опытов заключалась в том, что собака при показе сигнального цвета, который выполнял роль цветовой команды, совершала достаточно сложные движения. Так, B. Л. Дуров (1924) демонстрировал способность немецкой овчарки при показе одного цвета пройти в одном направлении, при показе второго цвета – в другом направлении и т. д. В этой ситуации собака ошибалась чаще, чем при подаче предмета требуемого цвета, но преимущественно выбирала направление правильно. Продолжая свой опыт, А. А. Бабушкин отобрал 3 собак (из 10 исходных) с наилучшим различением цветов и прошедших общий курс дрессировки. Он научил их выполнять знакомые им команды: «Сидеть!», «Стоять!» и «Лежать!» при показе соответственно красной, зеленой и синей карточек. Число правильных выполнений команд составило в среднем 73,8 % (немецкая овчарка), 77,6 % (кокер‑спаниель) и 86,7 % (доберман).
На обложке изображено выполнение этих навыков. Причем представлено в виде обычного двухфазного способа выработки условного рефлекса. Сначала подкрепляющий ранее усвоенный сигнал (голосовая команда) подается одновременно с обучающей командой (цветная карточка) или сразу после нее. Затем подкрепляющий сигнал не используется, и собака выполняет навыки только по предъявлении соответствующих им цветных карточек.
Несмотря на, казалось бы, подтверждение наличия цветового зрения у собак, остаются сомнения в их способности различать цвета. Так, С. А. Корытин (1984) считает, что собаки не различают цвета, а реагируют лишь на степень яркости. Наиболее одиозным примером служит высказывание финского зоолога и кинолога Ё. Бергмана (1992) по поводу результатов его соотечественницы A. Rosengren (1969): «Тот факт, что кокер‑спаниели могли различать цвета, еще не подтверждает этой способности у других пород». Странность такого заявления заключается в том, что за 80 с лишним лет до работы A. Rosengren ни разу не была замечена породная специфичность цветового зрения. А вот высказывание американского ветеринара и кинолога Л. Ф. Уитни (1997): «Насколько можно судить по полученным учеными данным, цвета собаки не различают. У них не существует разницы между яркостью и собственно цветом. Одна группа ученых заключила, завершив исследования, что собаки не различают цвета, и если вообще реагируют на цветовые стимулы, то весьма слабо». Примерно этого же мнения придерживается его соотечественница Е. X. Вайтли (1997). Интересно, что все перечисленные авторы самостоятельно не изучали этот вопрос. Они использовали единственное обоснование, сформулированное еще Е. М. Smith (1912) в объяснение неоднозначных результатов своей работы. Она предположила, что различение цветов едва ли может играть ориентирующую роль в нормальной жизни собак. С ней согласился Л. А. Орбели (1913). Уже наш современник Ё. Бергман (1992), допуская вероятность восприятия собаками цвета, отметил, что он мало что для них значит и в их жизни не играет большой роли. Однако работа В. К. Шепелевой (1954) показала, что подобное мнение, по крайней мере, не универсально. Ее дальнейший эксперимент заключался в следующем. С зеленым клапаном, закрывавшим вход в камеру, где две собаки получали подкормку, соединили тампон, смоченный в камфаре. После многократных таких сочетаний (цвет‑запах) тампон перенесли на красный клапан, закрывавший вход в камеру, где подкормку не давали. При многократных повторениях собаки ориентировались только на зеленый клапан. То же самое происходило при замене камфары тополевым маслом в сочетании с зеленым клапаном. А дальше создали, казалось бы, совсем простую «обонятельную ситуацию». Тампон с тополевым маслом соединили сначала с одним из пяти красных, потом с одним из пяти зеленых клапанов (обычно входы в камеры закрывали клапаны разных цветов). Собаки проявили хаотическую реакцию. И в первое время число правильных подходов (на запах) составляло 10…30 %. Интересно, что и в этих ситуациях проявились индивидуальные особенности животных. Так что в некоторых ситуациях цвет для собак важнее, чем даже запах, который принято считать главным для них раздражителем. А отмечаемая всеми исследователями слабость цветового зрения объясняется в первую очередь спецификой морфофизиологических характеристик ее глаз. Был сделан вывод: собаки обладают цветовым зрением. Однако условный рефлекс на цветовой тон образуется у них чрезвычайно медленно, еще более затрудняясь при изменении его интенсивности. Но образовавшиеся временные связи на цветовой тон достаточно прочные. Собаки с трудом переключаются на ориентирование при помощи обонятельного или двигательного анализатора. Обобщая информацию о способности собак различать цвета, уместно привести высказывание В. К. Шепелевой (1971) после успешного изучения цветового зрения у лесного хорька по той же методике, что и на собаках: «Видимо, распространенное среди экспериментаторов, изучающих наличие цветового зрения, мнение о весьма ограниченном числе видов животных, обладающих цветовым зрением, нельзя считать достаточно обоснованным ввиду крайне ограниченного количества соответствующих экспериментальных исследований и из‑за ошибок в методических приемах. Невозможно, чтобы среди обширного класса млекопитающих со столь высокоразвитыми органами зрения и тонкой организацией нервной системы цветовое зрение имелось бы только у приматов и эволюция не шла бы по линии выработки такой значимой для существования животных способности, как различение цветов». Следует заметить, что во всех работах по изучению цветового зрения даже не упоминались морфофизиологические особенности глаза собаки, хотя понятно, что именно они в первую очередь определяют цветовое зрение[4]. А ведь морфология сетчатки изучалась с середины XIX в., фактически одновременно с изучением цветового зрения. И даже в работах, посвященных колбочкам, фактическим цветоприемникам, их описывают в основном как часть сетчатки и менее как элемент глаза, обусловливающий остроту зрения. Поэтому сопоставим морфофизиологические характеристики глаза собаки с особенностями ее цветового зрения, т. е. попытаемся выяснить, почему оно такое, а не иное. Начнем с того, что в месте наилучшего видения (в центре овальной полосы) колбочек очень мало и они соединены по несколько штук с одной ганглиозной клеткой. Кроме того, они перемешаны с палочками (нецветоприемниками), которых даже здесь в 10 раз больше, чем колбочек, а на периферии сетчатки разница еще больше. Все эти факторы при низкой светочувствительности колбочек обусловливают слабое и нечеткое восприятие цветов, затрудняют их различение. Но и это еще не все «оппоненты» цветового зрения. Панорамность зрения в сочетании с преобладанием палочек, имеющих высокую светочувствительность, позволяет собаке даже при характерной для нее слабой дивергенции оптических осей одинаково хорошо видеть спереди и по сторонам, замечать малейшие изменения обстановки и различия в форме, размерах, удаленности, освещенности, особенно подвижности и других нецветовых признаков объектов. Все это, конечно, отвлекает собаку от цвета и препятствует адекватному ответу на него. Незнание или неучет мешающих факторов в сочетании с непродолжительностью тренировки были причиной некоторых неудачных работ по изучению цветового зрения у собак [8, 115]. Однако у известного дрессировщика В. Л. Дурова собака при общении с ним отдавала предпочтение цвету предмета, а не форме. Например, при показе круглой фарфоровой зеленой сахарницы она приносила зеленые мячи, а при показе круглой лакированной коричнево‑красной розетки приносила коричневые мячи. Причем последние были разного тона и лежали все вместе [60]. Приведенные свойства сетчатки глаза, конечно, обусловливают слабость цветового зрения, если колбочки охарактеризованы только с количественной стороны. А есть ли в них что‑то, свойственное именно собакам, и какое это имеет значение? Были выявлены два типа колбочек с использованием антител к их наружным сегментам (К. Gropp et al., 1993) и с помощью ЭРГ‑мигающей фотометрии (J. Neitz, G. H. Jacobs et al., 1989; G. H. Jacobs, 1993). Одни колбочки максимально чувствительны к свету с длиной волны 429…435 нм (для людей – фиолетовый цвет), другие колбочки – к свету с длиной волны около 555 нм (для людей желто‑зеленый цвет). И хотя неизвестно, воспринимают ли собаки эти цвета, как люди, исследователи предположили, что видимый спектр у собак делится на два оттенка – фиолетовый и сине‑фиолетовый (430…475 нм), который они, вероятно, видят как синий, и зеленовато‑желтый, желтый и красный (500…620 нм), который они, вероятно, видят как желтый. Собаки, наверное, воспринимают узкую часть видимого спектра как бесцветную. Цвета с двух концов спектра (синий и желтый), вероятно, воспринимаются ими наиболее насыщенными, а цвета промежуточных волн – менее интенсивными, как если бы они были смешаны с белым или серым. Поэтому в противоположность человеку, который обладает трихроматическим зрением и воспринимает волны видимого спектра как сотни отдельных цветов, цветовое зрение собак дихроматично со спектрально нейтральной зоной. На основании обнаруженной спектральной чувствительности колбочек американские исследователи предположили, какие цвета видят собаки. Исходя из максимальной чувствительности одного из типов колбочек в средней части спектра был сделан вывод о неспособности собак различать свет средних и длинных волн (J. Neitz et al, 1989). Для подтверждения проводили сравнение их с людьми (дальтониками), у которых отсутствуют колбочки, воспринимающие зеленый цвет, и потому не различающих красный и зеленый цвета (красно‑зеленая слепота). Во‑первых, колбочки, максимально чувствительные к зеленому цвету, воспринимают и красный (620 нм), но в меньшей степени, чем зеленый. Во‑вторых, и самое главное, многочисленными работами показана способность собак различать любые цвета, в том числе красный, любой яркости при соблюдении условий, соответствующих особенностям их цветового зрения. Возможно, на основании этих данных Е. X. Вайтли (1997) пишет, что, вероятно, у собак способность видеть цвет главным образом в спектре длинных волн света (красные оттенки) ограничена. Еще менее обоснованным представляется сформированное значительно раньше мнение, что собаки цветные изображения видят в серых тонах: желтый цвет воспринимается ими как светло‑серый, оранжевый и синий – более темным, красный, зеленый и фиолетовый – как темно‑серый [48]. Или что собаки видят большинство объектов серыми, будто полинявшими [12]. В целом произошла подмена понятия «различение цветов» понятием «видение цветов». Так вот, во всех работах изучали, как собаки различают цвета, а не какими они их видят. Можно выявить спектр поглощения света колбочками собаки, можно показать различение ею цветов, но нельзя сказать, каким каждый цвет видит собака (в нашем восприятии). Очень кратко это объяснил Г. И. Азимов (1958): «Животное смотрит глазами, а видит мозгом». Помимо особенностей нервной системы, анатомо‑физиологические характеристики глаз обусловливают еще одно свойство цветового зрения – индивидуальные особенности. Одной из причин может быть отклонение морфологии сетчатки от ее классической схемы – двухэтажной светло‑темной структуры. Светлая, тапетальная, т. е. отражающая, зона может быть незначительной по площади вплоть до ее отсутствия за счет расширения темной, нетапетальной, т. е., наоборот, поглощающей зоны. Может быть и обратная картина, вплоть до отсутствия пигментного слоя (частичный альбинизм). Разумеется, в глазу создаются разные световые условия в связи с отражением, рассеиванием и поглощением света. В результате меняется эффективность работы колбочек как цветоприемников. Да и сами колбочки могут отличаться по набору зрительных пигментов, что неминуемо отражается на разном восприятии цветов у отдельных собак. Индивидуальные особенности цветового зрения могут обусловливаться также расширением панорамности зрения по мере смещения расположения глаз от осевой линии головы на ее боковые поверхности, т. е. расширением монокулярного зрения (крайние варианты: борзые, колли, лайки). Исходя из большей возможности вести боковой обзор, у собак этого типа более вероятно отвлечение на посторонние факторы во время отработки какого‑то действия в связи с цветом по сравнению с собаками с противоположным расположением глаз – около осевой линии головы (крайние варианты: боксер, бульдоги, мопс, пекинес). Таким образом, при общении с собакой следует использовать и цветовое зрение. Вероятно, в связи с цветом объекта у них можно вырабатывать не только простые условные рефлексы, но и более сложные формы поведения – навыки. Правда, при этом необходимо учитывать все особенности цветового зрения. Оно проявляется значительно слабее и менее обеспечивает соответствующую память, чем восприятие других характеристик объекта: размера, формы, удаленности и особенно подвижности. Решающую роль может сыграть и окружающая обстановка. Значит, малейшие изменения, даже незаметные для человека, могут отвлечь животное, помешать сосредоточиться именно на цвете. Следовательно, выработка каких‑либо навыков у собаки в связи с цветом объекта требует многократных повторов при соблюдении неизменности других характеристик объекта и окружающей обстановки. Учитывая непрочную цветовую память животного, эти навыки придется периодически закреплять. С другой стороны, цвет объекта, в том числе детали одежды, может запомниться собакой независимо от воли и даже просто внимания общающегося с ней человека
|
||
|
|
Последнее изменение этой страницы: 2021-01-14; просмотров: 167; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.119 (0.017 с.) |