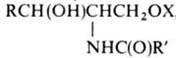Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Химизм распада глюкозы в аэробных условиях.Содержание книги
Поиск на нашем сайте
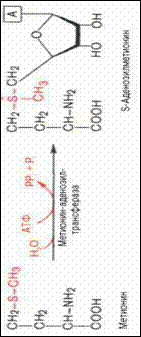
Биосинтез желчных кислот. Первичные желчные кислоты возникают в гепатоцитах из холестерина. При образовании холиевой кислоты происходит восстановление двойной связи, окисление атомов 7 и 12 концевой метильной группы боковой цепи холестерина с последующим отщеплением пропионовой кислоты. Вторичные жел/кислоты образуютсяв кишечнике из первичных путемчастичного восстановления последних под влиянием микроорганизмов. Желчные кислоты способствуют эмульгированию пищевых жиров, участвуют наряду с колипазой в активировании пенкреатической липазы(сдвигая ее рН в кислую сторону), обеспечивают всасывание продуктов переваривания липидов. S-аденозилметионин, Фактическим донором метильных групп в реакциях трансметилирования является не свободный метионин, а так называемый активный метионин – S-аденозилметионин, который образуется в процессе АТФ-зависимой реакции, катализируемой метионин-аденозилтрансферазой. . Своеобразие данной реакции заключается в том, что СН3-группа ме-тионина активируется под действием положительного заряда соседнего атома серы. S-аденозилметионин участвует во всех реакциях, где метильная группа используется в биосинтетических реакциях: например, в синтезе адреналина, креатинина, тимина, фосфатидилхолина, бетаина и др. Распад пуриновых оснований Конечные продукты: мочевая кислота, СО2, Н2О,фосфаты, аммиак. 108. Распад пиримидин. Осн.: Продукты распада:аммиак, СО2, Н2О,фосфаты Обезвреживание аммиака. Основная масса аммиака обезвреживается в печени с образованием мочвины и в почках с образованием аммонийных солей. Мочевина и соли аммония это конечные продукты азотистого обмена, они выводятся почками. Способы: _основной реакцией выведения аммиака является синтез глутамина под действием глутаминсинтетазы. Глутамат+NН3+АТФ->Глутамин +АДФ+Pi. Глутаминсинтетаза обладает высоким сродством к аммиаку и благодаря этой реакции в крови и в тканях поддерживается низкая концентрация NН3. Глутамин явл транспортной формой аммиака т.к. представляет собой нейтральную амино к-ту, способную протекать через клеточные мембраны путем облегченной диффузии.Образовавшийся в тканях глутамин транспортируется в почки и кишечник. В клетках киш от глутамина отщепляется амидная группа в виде NН3, образовавшийся глутамат трансаминируется с образованием аммиака: Глутамин+Н2О à глутамат+ NН3
Глутамат +пируватà альфа-кетоглутамат=аланин. Первая р-я катализируется глутаминазой, вторая - АЛТ -н.сист: аммиак выводится при помощи восстановительного аминирования, альфа-кетоглутарата под действием глутаматдегидрогеназы, Глутамат в дальн-м может превращаться в глутамин. NН3 Альфа-кетоглутара à глутамат NН3 ГД à глутамин Глутаминсинтетаза
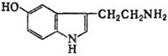
-из мышц избыток азота выводится в виде аланина. Аланин получается из ПВК трансаминированием. Источником ПВК в м-цах является глю и также распадающиеся аминокислоты. Аланин из мышц поступает в печень, где из него непрямым дезаминированием получается амиак и ПВК. Аммиак обезвреживается, а ПВК включается в глюконеогенез, образовавшаяся в результате глюконеогенеза глю из печени поступает в ткани, а там окисляется до пирувата. 110.Синтез серотонина В организме человека и животных С. синтезируется во мн. тканях из триптофана путем его гидроксилирования (фермент триптофангидрок-силаза) с образованием 5-гид-рокси-триптофана, кот. затем под действием декарбокси-лазы ароматич. L-аминокислот превращается в С. Играет важную роль в деятельности центр. и периферич. нервной системы в качестве медиатора (передатчика) нервных импульсов. 111. гормоны задней доли гипофиза: Вазопрессин(В) и окситоцин(О) синтезируются в рибосомах. В гипоталамусе синтезируются белки нейрофизин I, II и III способные связывать В иО и транспортировать их в нейросекреторные гранулы гипоталамуса, из ядер гипоталамуса мигрируют вдоль аксона и достигают задней доли гипофиза – резерв гормонов Химич.строние:О: S----------------------------S Н-Цис-Тир-Иле-Глн-Асн-Цис-Про-Лей-Гли-СО-NН2 S---------------------------S Н-Цис-Тир-Иле-Глн-Асн-Цис-Про-Арг-Гли-СО-NН2 Физиолог.значение: О: сокращение гладких мышц матки и циркулярных параальвеолярных мыш волокон мол.желез. В: стимуляция сокращения гл.мышечн.волокон сосудов, регуляция водного обмена. 112. Гормоны передней доли гипофиза: Гормон роста(СТГ), кортикотро-пин, тиротропин, пролактин, фолликулостимулирующий, лютеинизирующий
СТГ под его действием усиливается синтез белка, ДНК, РНК и гликогена, мобилизация жиров из депо и распад высших жирных кислот и глюкозы в тканях. + обладает лактогенной активностью. Кортикотропин:: стимуляция синтеза и секреции гомонов коры надпочечников. Пролактин: стимуляция развития мол.желез и лактации Тиротропин: контролирование развития и функции щит.жел, регуляция и синтез в кровь тиреоидных гормонов.. Горм корк в-ва надпоч. гормоны коркового вещества надпочечников условно делят на глюкокортикоиды (корти-костероиды, оказывающие влияние на обмен углеводов, белков, жиров и нуклеиновых кислот) и минералокортикоиды (кортикостероиды, оказывающие преимущественное влияние на обмен солей и воды). К первым относятся кортикостерон, кортизон, гидрокортизон (кортизол), 11-дезоксикортизол и 11-дегидрокортикостерон, ко вторым – дезоксикортико-стерон и альдостерон. Установлено, что предшественником кортикостероидов является холес-терин(ол) и процесс стероидогенеза, как и нормальное гистологическое строение и масса надпочечников, регулируется АКТГ гипофиза. В свою очередь синтез АКТГ в гипофизе, а значит, и кортикостероидов в корковом веществе надпочечников регулируется гипоталамусом, который в ответ на стрессовые ситуации секретирует кортиколиберин. Глюкокортикоиды оказывают разностороннее влияние на обмен веществ в разных тканях. В мышечной, лимфатической, соединительной и жировой тканях глюкокортикоиды, проявляя катаболическое действие, вызывают снижение проницаемости клеточных мембран и соответственно торможение поглощения глюкозы и аминокислот; в то же время в печени они оказывают противоположное действие. Конечным итогом воздействия глюкокорти-коидов является развитие гипергликемии, обусловленной главным образом глюконеогенезом. Минералокортикоиды (дезоксикортикостерон и альдостерон) регулируют главным образом обмен натрия, калия, хлора и воды; они способствуют удержанию ионов натрия и хлора в организме и выведению с мочой ионов калия. По-видимому, происходит обратное всасывание ионов натрия и хлора в канальцах почек в обмен на выведение других продуктов обмена, в частности мочевины. 115.Гормоны пептидной природы: Пептидные и белковые гормоны включают от 3 до 250 и более аминокислотных остатков. Это гормоны гипоталамуса и гипофиза (тироли-берин, соматолиберин, соматостатин, гормон роста, кортикотропин, тире-отропин и др. – см. далее), а также гормоны поджелудочной железы (инсулин, глюкагон). Могут вызвать быструю ответную реакцию орг. Путемувеличения активности уже сущ-х в тканях ферментов. 116. Релизинг-факторы: Под влияниемразличных раздражителей возникают импульсы в специализированных рецепторах, и направл в ЦНС. В гипоталамусе в ответ синтезируются биол.активн.гормональные в-ва-релизинг-факторы. Которые через портальную систему сосудов достигают гормонсинтезирующих клетокгипофиза, изменяя их активность стимулируя или тормозя. 117.гормоны ит.железы: Тироксин, трийодтирамин и кальцитонин.
Биосинтез тироксина происходит в фолликулах щитовидной железы путем конденсации двух остатков молекул дииодтирози-на, входящих в состав тиреоглобулина - гликопротеина, содержащего ок. 5 тыс. аминокислотных остатков (из них 120-остатки тирозина). Иодирование остатков тирозина осуществляется иодом, к-рый образуется путем ферментативного окисления иодидов, поступающих в щитовидную железу вместе с кровью. Механизм биосинтеза Т., по-видимому, включает окисление остатка дииодтирозина в тирео-глобулине до своб. радикала. Образующиеся в результате синтеза Т. остатки пировиноградной к-ты или серина остаются в составе молекулы тиреоглобулина.
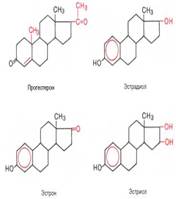
У человека и высших животных он усиливает энергетич. обмен (в т.ч. поглощение О2 тканями, увеличение теплопродукции), влияет на рост и дифференцировку тканей, стимулирует сердечную деятельность, повышает возбудимость нервной системы. У земноводных и нек-рых костистых рыб стимулирует метаморфоз. В основе механизма физиол. действия Т. лежит его взаимод. со специфич. рецепторами клеточных ядер и регулирующее влияние на процессы синтеза РНК и белка. Ингсулин человека состоит из двух пептидных цепей (А и В), соединенных двумя ди-сульфидными мостиками; третий дисульфидный мостик находится в цепи А биосинтез инсулина осуществляется в β-клетках панкреатических островков из своего предшественника проинсулина В физиологической регуляции синтеза инсулина доминирующую роль играет концентрация глюкозы в крови. Так, повышение содержания глюкозы в крови вызывает увеличение секреции инсулина в панкреатических островках, а снижение ее содержания, наоборот,– замедление секреции инсулина. Этот феномен контроля по типу обратной связи рассматривается как один из важнейших механизмов регуляции содержания глюкозы в крови. На секрецию инсулина оказывают влияние, кроме того, электролиты (особенно ионы кальция), аминокислоты, глюкагон и секретин. Приводятся доказательства роли циклазной системы в секреции инсулина. Предполагают, что глюкоза действует в качестве сигнала для активирования аденилат-циклазы, а образовавшийся в этой системе цАМФ – в качестве сигнала для секреции инсулина. Эстрогены Основным местом синтеза женских половых гормонов – эстрогенов (от греч. oistros – страстное влечение) – являются яичники и желтое тело; доказано также образование этих гормонов в надпочечниках, семенниках и плаценте.
Основная биологическая роль эстрогенов и прогестерона, синтез которых начинается после наступления половой зрелости, заключается в обеспечении репродуктивной функции организма женщины. В этот период они вызывают развитие вторичных половых признаков и создают оптимальные условия, обеспечивающие возможность оплодотворения яйцеклетки после овуляции. Прогестерон выполняет в организме ряд специфических функций: подготавливает слизистую оболочку матки к успешной имплантации яйцеклетки в случае ее оплодотворения, а при наступлении беременности основная роль – сохранение беременности; оказывает тормозящее влияние на овуляцию и стимулирует развитие ткани молочной железы. Эстрогены оказывают анаболическое действие на организм, стимулируя синтез белка.
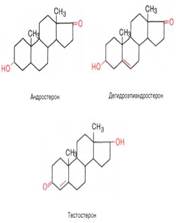
Андрогены
Биологическая роль андрогенов в мужском организме в основном связана с дифференцировкой и функционированием репродуктивной системы, причем в отличие от эстрогенов андрогенные гормоны уже в эмбриональном периоде оказывают существенное влияние на дифференцировку мужских половых желез, а также других тканей, определяя характер секреции гонадотропных гормонов у взрослых. Во взрослом организме андрогены регулируют развитие мужских вторичных половых признаков, сперматогенез в семенниках и т.д. Следует отметить, что андрогены оказывают значительное анаболическое действие, выражающееся в стимуляции синтеза белка во всех тканях, но в большей степени в мышцах. Пргестерон Женский стероидный гормон позвоночных, вырабатываемый желтым телом яичника, плацентой (в меньших количествах – семенниками и корой надпочечников) Прогестерон выполняет в организме ряд специфических функций: подготавливает слизистую оболочку матки к успешной имплантации яйцеклетки в случае ее оплодотворения, а при наступлении беременности основная роль – сохранение беременности; оказывает тормозящее влияние на овуляцию и стимулирует развитие ткани молочной железы. Эстрогены оказывают анаболическое действие на организм, стимулируя синтез белка. 123.Гестагены: (прогестины, гормоны желтого тела), гормоны регулирующие беременность у плацентарных животных. 124. Гормоны оказывают действие на специализированные органы и ткани (органы- мишени) Вызывая ответные реакции на действие раздражителей. Химизм распада глюкозы в аэробных условиях.
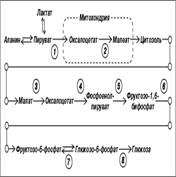
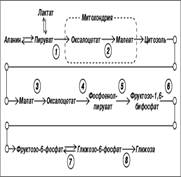
Энергетический выход=1900кДж/ моль глюкозы аккумулируется в молекулах и м.б. использовано работающими механизмами клетки. 66. Глюконеогенез – синтез глюкозы из неуглеводных предшественников: основные- пируват и лактат, промежуточные – глюкогенные аминокислоты и глицерин. Цикл Кори Начинается с образования лактата в мышцах в результате анаэробного гликолиза..Лактат переносится кровью в печень, где в процессе глюконеогенеза превращается в глюкозу, которая затем с током крови может возвращаться в работающую мышцу. Итак печень снабжает мышцу глюкозой и, следовательно, энергией для сокращений. В печени часть лактата может окисляться до СО2 и Н2О, превращаясь в пируват и далее в общих путях катаболизма.
67-68. Пентозофосфатный путь превращения глюкозы. Представлен двумя последовате-льными ветвями: окислительной и неокислительной. Окислительная ветвь: 1) глюкозо-6-Ф + НАДФ -> 6фос-фоглюконолактон + НАДФ*Н (фермент-дегидрогеназа) 2)6фосфоглюконолактон + Н2О -> 6фосфоглюконат + Н (фермент – лактоназа)
3) 6-фосфоглюконат + НАДФ -> рибозо-5-фосфат + СО2 + НАДФ*Н (фермент – фосфоглюконатдегидрогеназа) Неокислительная ветвь: 1)рибозо-5-фосфат->рибозо-5-фо-сфат (пентозофосфатизомераза) 2)рибозо-5-фосфат -> ксилулозо-5-фосфат (пентозофосфатэпимераза) 3)ксилулозо-5-фосфат + рибозо-5-фосфат -> седогептулозо-7-фосфат + глицеральдегид-3-фосфат (транскетолаза) 4) седогептулозо-7-фосфат + глицеральдегид-3-фосфат -> фруктозо-6-фосфат + эритрозо-4-фосфат (транскетолаза) 5) ксилулозо-5-фосфат + эритрозо-4-фосфат -> фруктозо-6-фосфат + глицеральдегид-3-фосфат(транскетолаза) Биологич.роль: образование НАДФ*Н2, синтез рибозо-5-фосфата, который используется в реакциях синтеза РНК, ДНК, АТФ, НАД, ФАД, КоА. 69. Гликоген – депонированная форма глюкозы
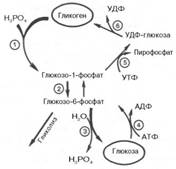
Жирными стрелками указан путь распада, тонкими - путь синтеза. Цифрами обозначены ферменты: 1 - фосфорилаза; 2 - фос-фоглюкомутаза; 3 - глюкозо-6-фосфатаза; 4 - гексокиназа (глюкокиназа); 5 - глюко-зо-1-фосфат-уридилтрансфераза; 6 - глико-генсинтаза. Можно считать, что сохранение постоянства концентрации глюкозы в крови является результатом одновременного протекания двух процессов: поступления глюкозы в кровь из печени и потребления ее из крови тканями, где она используется в первую очередь как энергетический материал. 70. Гликогенозы - Ряд наследственных болезней связан с нарушением обмена гликогена. Эти болезни получили название гликогенозов. Они возникают в связи с дефицитом или полным отсутствием ферментов, катализирующих процессы распада или синтеза гликогена, и характеризуются избыточным его накоплением в различных органах и тканях. Гликогеноз I типа (болезнь Гирке) встречается наиболее часто, обусловлен наследственным дефектом синтеза фермента глюкозо-6-фосфатазы в печени и почках. 71. Взаимопревращение углево-дов. Неиспользованные углеводы, их фосфорилированные формы, получающиеся в результате превращения в пентозном цикле Iстадии, могут взаимодействовать между собой с образованием, как новых продуктов, ичпользующихся в различных синтетических процессах, так и неиспользованные пентозы которые могут вновь превращаться в гексозы => в конце в глюкозо-6-фосфат. 72. Глюконеогенез – синтез глюкозы из неуглеводных предшественников: основные- пируват и лактат, промежуточные – глюкогенные аминокислоты и глицерин. Цикл Кори Начинается с образования лактата в мышцах в результате анаэробного гликолиза..Лактат переносится кровью в печень, где в процессе глюконеогенеза превращается в глюкозу, которая затем с током крови может возвращаться в работающую мышцу. Итак печень снабжает мышцу глюкозой и, следовательно, энергией для сокращений. В печени часть лактата может окисляться до СО2 и Н2О, превращаясь в пируват и далее в общих путях катаболизма.
73.переваривание жиров. У взрослых неэмульгированные триглицериды, составляющие основную массу пищевого жира, проходят через желудок без особых изменений.После того как химус попадает в двенадцатиперстную кишку Одновременно начинается эмульгирование жира. Наиболее мощное эмульгирующее действие на жиры оказывают соли желчных кислот, попадающие в двенадцатиперстную кишку с желчью в виде натриевых солей. Большая часть желчных кислот конъюгирована с глицином или таурином. По химической природе желчные кислоты являются производными холановой кислоты. Желчные кислоты способствуют эмульгированию пищевых жиров, участвуют наряду с колипазой в активации панкреатической липазы (сдвигая ее рН в кислую сторону) обеспечивают переваривание продуктов переваривания липидов. 74. Ресинтез триацилглице-ринов в стенке кишечника. Процесс синтеза жира в энтероцитах из компонентов мицелл называется ресинтезом жира. В процессе ресинтеза происходит образование жиров, близких по составу к жирам организма. При помощи хиломикронов осуществляется доставка пищевого (экзогенного) жира из кишечника в другие ткани (главным образом в жировую ткань. В жировой ткани из продуктов гидролиза триацилглицеринов снова происходит ресинтез жира (второй), и он депонируется там, пока не будет востребован. Аполипопротеины- это апобелки входящие в состав хиломикронов Апобелки выполняют не только структурную функцию, но и обеспечивают активное участие комплексов ЛП в транспорте липидов в токе крови от мест их синтеза к клеткам периферических тканей, а также обратный транспорт холестерина в печень для дальнейших метаболических превращений. Апобелки выполняют функцию лигандов во взаимодействии ЛП со специфическими рецепторами на клеточных мембранах, регулируя тем самым гомеостаз холестерина в клетках и в организме в целом. 75. Липиды – вещества, нерастворимые в воде, но растворяющиеся в органических растворителях, содержащие в своих молекулах высшие углеводородные радикалы. Классификация: -простые (жиры, воска) -сложн (фосфо- и гликолипиды) -изопреноиды (стероиды, каротиноиди) Л.участвуют в построении биологических мембран, являются важнейшим источником энеогии (триацилглицерины), обеспечивают организм витF, холестерин явл предшественником желчных кислот, стероидных гормонов, витД3, Л.выполняют защитную функц. 76. Классификация фосфоли-пидов: глицерофосфолипиды (в роли спирта выступает глицерол), и сфинголипиды (в роли спирта – сфингозин) Биосинтез фосфолипидов триглицериды и фосфатидные синтезируются на основе глицерофосфата. Эти соединения образуются из глицерина в результате переаминирования с АТФ (катализатор- глицеро-киназа) или диоксиацетонфосфата (образуется в ходе гликолиза) в результате восстановления за счет НАДФ*Н2, катализируемого глицерофосфат дегидрогеназой. Глицерофосфат реагирует с двумя молекулами ацил-КоА, образуя фосфатидные кислоты. Фосфатидная кислота под действием фосфатазы теряет остаток фосфата. Высвободившийся диглицерид реагирует с третьей молекулой ацил-КоА, образуя триглицерид. Фосфолипиды играют важную роль в структуре и функции клеточных мембран, активации мембранных и лизосомальных ферментов, в проведении нервных импульсов, свертывании крови, иммунологических реакциях, процессах клеточной пролиферации и регенерации тканей, в переносе электронов в цепи «дыхательных» ферментов. Особая роль фосфолипидам отводится в формировании липо-протеидных комплексов. 77. Классификация сфинголи-пидов: фосфосфинголилиды и гликосфинголипиды. Сфинголипиды уч-ют в передаче клеточных сигналов (нервная ткань) СН3-(СН2)12-СН=СН-СН-СН-NН-С-R ОН СН2 О О Р О О-СН2-СН2 N(СН3)3 Холин 78. Ганглиозиды — представи-тели наиболее сложно построенных гликолипидов. Они представляют большое семейство мембранных липидов, выполняющих, по-видимому, рецепторные функции. Характерной особенностью ганглиозидов является наличие остатков N-ацетилнейраминовой кислоты
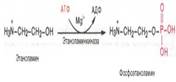
Г. широко распространены в тканях мозга 79. Биосинтез кефалина. Первоначально этаноламин при участии соответствующей киназы фосфорилируется с образованием фосфо-этаноламина:
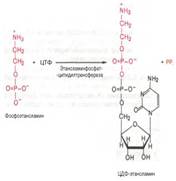
Затем фосфоэтаноламин взаимодействует с ЦТФ, в результате чего образуются цитидиндифосфатэтаноламин (ЦДФ-этаноламин) и пирофос-фат (PPi):
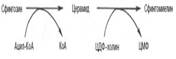
В следующей реакции ЦДФ-этаноламин, взаимодействуя с 1,2-дигли-церидом, образующим-ся при дефосфорилировании фосфатидной кислоты, превраща-ется в фосфатидилэтаноламин. Реакция катализируется фермен-том этаноламинфосфотрансфер-азой: ЦДФ-этаноламин + 1,2-диглицерид –> Фосфати-дилэтаноламин + ЦМФ Синтез лецитина: 80. Биосинтез церамида: Интермедиатом в биосинтезе сфингомиелина является церамид (N-ацилсфингозин), который образуется при взаимодействии сфингозина с ацил-КоА. Сфингомиелин синтезируется в результате взаимодействия (реакции) церамида с ЦДФ-холином:
|
|||||||||
|
|
Последнее изменение этой страницы: 2021-01-09; просмотров: 71; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.139.234.68 (0.016 с.) |