Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Цитоархитектоника и миелоархитектоника новой корыСодержание книги
Поиск на нашем сайте
В соответствии с делением полушарий на пять долей выделяют пять основных областей – лобную, теменную, височную, затылочную и островковую, имеющие различия в строении и выполняющие разные функции. Однако общий план строения новой коры одинаков. Новая кора – это слоистая структура (рис. 37). I - молекулярный слой, образован преимущественно нервными волокнами, идущими параллельно поверхности. Среди параллельных волокон расположено небольшое количество зернистых клеток. Под молекулярным слоем расположен II слой – наружный зернистый. III слой – наружный пирамидный, IV слой, внутренний зернистый, V слой – внутренний пирамидный и VI слой – мультиформный. Название слоев дано по названию нейронов. Соответственно, во II и IV слоях – сомы нейронов имеют округлую форму (клетки-зерна) (наружный и внутренний зернистый слои), а в III и IV слоях сомы имеют пирамидную форму (в наружном пирамидном – малые пирамиды, а во внутреннем пирамидном – большие пирамиды или клетки Беца). VI слой характеризуется наличием нейронов разнообразной формы (веретенообразной, треугольной и др.). Главные афферентные входы в кору полушарий большого мозга – нервные волокна, идущие из таламуса. Корковые нейроны, которые воспринимают афферентные импульсы, идущие по этим волокнам, называют сенсорными, а зона, где расположены сенсорные нейроны – проекционными зонами коры. Главные эфферентные выходы из коры – аксоны пирамид V слоя. Это эфферентные, двигательные нейроны, участвующие в регуляции двигательных функций. Большинство нейронов коры – вставочные, участвующие в переработке информации и обеспечивающие межкортикальные связи.
Типичные нейроны коры
Римскими цифрами обозначены клеточные слои I - молекулярный сдой; II - наружный зернистый слой; III - наружный пирамидный слой; IV - внутренний зернистый слой; V - внутренний приамидный слой; VI-мультиформный слой. а - афферентные волокна; б - типы клеток, выявляемые на препаратах, импрегнированных по методу Голдбжи; в - цитоархитектоника, выявляемая при окрашивании по Нисслю. 1 - горизонтальные клетки, 2 - полоска Кеса, 3 - пирамидные клетки, 4 – звездчатые клетки, 5 - наружная полоска Белларже, 6 - внутренняя полоска Белларже, 7 - видоизмененная пирамидная клетка. Рис. 37. Цитоархитектоника (А) и миелоархитектоника (Б) коры полушарий большого мозга.
При сохранении общего плана строения было установлено, что разные участки коры (в пределах одной области) отличаются по толщине слоев. В некоторых слоях можно выделить несколько подслоев. Кроме того, имеются различия клеточного состава (разнообразие нейронов, плотность и их расположение). С учетом всех этих различий Бродман выделил 52 участка, которые назвал цитоархитектоническими полями и обозначил арабскими цифрами от 1 до 52 (рис.38 А,Б).
А вид сбоку. Б срединно-сагиттальный; срез. Рис. 38. Схема расположения полей по Бордману
Каждое цитоархитектоническое поле отличается не только клеточным строением, но и расположением нервных волокон, которые могут идти как в вертикальном, так и в горизонтальном направлениях. Скопление нервных волокон в пределах цитоархитектонического поля называют миелоархитектоникой. В настоящее время все большее признание находит “колончатый принцип” организации проекционных зон коры. Согласно этому принципу каждая проекционная зона состоит из большого количества вертикально ориентированных колонок, диаметром приблизительно в 1 мм. Каждая колонка объединяет около 100 нейронов, среди которых имеются сенсорные, вставочные и эфферентные нейроны, связанные между собой синаптическими связями. Одиночная “корковая колонка” участвует в переработке информации от ограниченного количества рецепторов, т.е. выполняет специфическую функцию. Система волокон полушарий
Оба полушария имеют три типа волокон. По проекционным волокнам возбуждение поступает в кору от рецепторов по специфическим проводящим путям. Ассоциативные волокна связывают между собой различные области одного и того же полушария. Например, затылочную область с височной, затылочную – с лобной, лобную – с теменной областью. Комиссуральные волокна связывают симметричные участки обоих полушарий. Среди комиссуральных волокон выделяют: переднюю, заднюю мозговые спайки и мозолистое тело (рис. 39 А.Б).
Рис. 39А. а - медиальная поверхность полушария; б - верхнеальтеральная поверхность полушария; А - лобный полюс; В - затылочный полюс; С - мозолистое тело; 1 - дугообразные волокна большого мозга соединяют между собой соседние извилины; 2 - пояс - пучок обонятельного мозга лежит под сводчатой извилиной, простирается из области обонятельного треугольника до крючка; 3 - нижний продольный пучок связывает затылочную и височную область; 4 - верхний продольный пучок связывает лобную, затылочную, височную доли и нижнетеменную дольку; 5 - крючковидный пучек расположен у переднего края островка и соединяет лобный полюс с височным.
Рис. 39Б. Кора головного мозга на поперечном разрезе. Оба полушария соединены пучками белого вещества, образующими мозолистое тело (комиссуральные волокна). Рис. 39. Схема ассоциативных волокон
Ретикулярная формация
Ретикулярная формация (сетчатое вещество мозга) была описана анатомами в конце прошлого века. Ретикулярная формация начинается в спинном мозге, где она представлена желатинозной субстанцией основания заднего мозга. Основная ее часть находится в центральном стволе мозга и в промежуточном мозге. Она состоит из нейронов различной формы и размеров, которые имеют обширные ветвящиеся отростки, идущие в разных направлениях. Среди отростков выделяют короткие и длинные нервные волокна. Короткие отростки обеспечивают локальные связи, длинные - формируют восходящие и нисходящие пути ретикулярной формации. Скопления нейронов образуют ядра, которые находятся на разных уровнях мозга (спинном, продолговатом, среднем, промежуточном). Большинство ядре ретикулярной формации не имеют четких морфологических границ и нейроны этих ядер объединяются только по функциональному признаку (дыхательный, сердечно-сосудистый центр и др.). Однако на уровне продолговатого мозга выделяют ядра с четко обозначенными границами - ретикулярное гигантоклеточное, ретикулярное мелкоклеточное и латеральное ядра. Ядра ретикулярной формации моста по существу являются продолжением ядер ретикулярной формации продолговатого мозга. Наиболее крупные из них - каудальное, медиальное и оральное ядра. Последнее переходит в клеточную группу ядер ретикулярной формации среднего мозга и ретикулярное ядро покрышки мозга. Клетки ретикулярной формации являются началом как восходящих, так и нисходящих путей, дающих многочисленные коллатерали (окончания), которые образуют синапсы на нейронах разных ядер центральной нервной системы. Волокна ретикулярных клеток, направляющихся в спинной мозг, образуют ретикулоспинальный тракт. Волокна восходящих трактов, начинающихся в спинном мозге, связывают ретикулярную формацию с мозжечком, средним мозгом, промежуточным мозгом и корой полушарий большого мозга. Выделяют специфические и неспецифические ретикулярной формации. Например, некоторая часть восходящих путей ретикулярной формации получают коллатерали от специфических путей (зрительных, слуховых и т.д.), по которым афферентные импульсы передаются в проекционные зоны коры. Неспецифические восходящие и нисходящие пути ретикулярной формации влияют на возбудимость различных отделов мозга, в первую очередь коры полушарий большого мозга и спинной мозг. Эти влияния по функциональному значению могут быть как активирующими, так и тормозными, поэтому выделяют: 1) восходящее активирующее влияние, 2) восходящее тормозное влияние, 3) нисходящее активирующее влияние, 4) нисходящее тормозное влияние. На основании этих факторов ретикулярную формацию рассматривают как регулирующую неспецифическую систему мозга. Наиболее изучено активирующее влияние ретикулярной формации на кору полушарий большого мозга. Большинство восходящих волокон ретикулярной формации диффузно оканчивается в коре полушарий и поддерживает ее тонус, обеспечивает внимание. Примером тормозных нисходящих влияний ретикулярной формации является снижение тонуса скелетных мышц человека во время определенных стадий сна. Нейроны ретикулярной формации чрезвычайно чувствительны к гуморальным веществам. Это опосредованный механизм влияния различных гуморальных факторов и эндокринной системы на высшие отделы мозга. Следовательно, тонические воздействия ретикулярной формации зависят от состояния всего организма (рис.40).
Рис. 40. Активирующая ретикулярная система (АРС) — нервная сеть, по которой сенсорное возбуждение передается от ретикулярной формации ствола мозга к неспецифическим ядрам таламуса. Волокна от этих ядер регулируют уровень активности коры. Подкорковые ядра
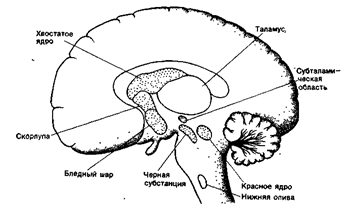
Подкорковые ядра входят в состав конечного мозга и расположены внутри белого вещества полушарий большого мозга. К ним относят хвостатое тело и скорлупу, объединяемые под общим названием “полосатое тело” (стриатум) и бледный шар, состоящий из чечевицеобразного тела, шелухи и миндалины. Подкорковые ядра и ядра среднего мозга (красное ядро и черная субстанция) составляют систему базальных ганглиев (ядер)(рис.41). К базальным ганглиям поступают импульсы от двигательной коры и мозжечка. В свою очередь, сигналы от базальных ганглиев направляются к двигательной коре, мозжечку и ретикулярной формации, т.е. существуют две нейронные петли: одна связывает базальные ганглии с двигательной корой, другая - с мозжечком.
Рис. 41. Система базальных ганглиев Подкорковые ядра принимают участие в регуляции двигательной активности, регулируя сложные движения при ходьбе, поддержании позы, при еде. Они организуют медленные движения (перешагивание через препятствия, вдевание нитки в иголку и т.д.). Имеются данные, что полосатое тело участвует в процессах запоминания двигательных программ, так как раздражение этой структуры приводит к нарушению обучения и памяти. Полосатое тело оказывает тормозное влияние на различные проявления двигательной активности и на эмоциональные компоненты двигательного поведения, в частности на агрессивные реакции. Основными медиаторами базальных ганглиев являются: дофамин (особенно в черной субстанции) и ацетилхолин. Поражение базальных ганглиев вызывает медленные извивающиеся непроизвольные движения, на фоне которых возникают резкие сокращения мышц. Непроизвольные порывистые движения головы и конечностей. Болезнь Паркинсона, основными симптомами которой является тремор (дрожание) и мышечная ригидность (резкое повышение тонуса мышц-разгибателей). Из-за ригидности больной с трудом может начать движение. Постоянный тремор препятствует выполнению мелких движений. Болезнь Паркинсона возникает при поражении черной субстанции. В норме черная субстанция оказывает тормозное влияние на хвостатое ядро, скорлупу и бледный шар. При ее разрушении тормозные влияния устраняются, в результате чего усиливается возбуждающее базальных ганглиев на кору головного мозга и ретикулярную формацию, что вызывает характерные симптомы болезни.
Лимбическая система
Лимбическая система представлена расположенными на границе отделами новой коры (неокортекса) и промежуточного мозга. Она объединяет комплексы структур разного филогенетического возраста, часть из которых является корковыми, а часть - ядерными. К корковым структурами лимбической системы относят гиппокампальную, парагиппокампальную и поясную извилины (старя кора). Древняя кора представлена обонятельной луковицей и обонятельными бугорками. Новая кора - часть лобной, островковой и височной коры. Ядерные структуры лимбической системы объединяют миндалину и септальные ядра и передние таламические ядра. Многие анатомы причисляют к лимбической системе преоптическую область гипоталамуса и маммилярные тела. Структуры лимбической системы образуют 2-х сторонние связи и связаны с другими отделами головного мозга. Лимбическая система контролирует эмоциональное поведение и регулирует эндогенные факторы, обеспечивающие мотивации. Положительные эмоции связаны преимущественно с возбуждением адренэргических нейронов, а отрицательные эмоции так же как страх и тревога - с недостатком возбуждения норадренэргических нейронов. Лимбическая система участвует в организации ориентировочно-исследовательского поведения. Так, в гиппокампе обнаружены нейроны “новизны”, меняющие свою импульсную активность при появлении новых раздражителей. Гиппокамп играет существенную роль в поддержании внутренней среды организма, участвует в процессах обучения и памяти. Следовательно, лимбическая система организует процессы саморегуляции поведения, эмоции, мотивации и памяти (рис.42).
Рис. 42. Лимбическая система Автономная нервная система
Автономная (вегетативная) нервная система обеспечивает регуляцию внутренних органов, усиливая или ослабляя их деятельность, осуществляет адаптивно-трофическую функцию, регулирует уровень метаболизма (обмен веществ) в органах и тканях (рис.43, 44).
1 - симпатический ствол; 2 - шейно-грудной (звездчатый) узел; 3 – средний шейный узел; 4 - верхний шейный узел; 5 - внутренняя сонная артерия; 6 - чревное сплетение; 7 - верхнее брыжечное сплетение; 8 - нижнее брыжечное сплетение Рис. 43. Симпатическая часть вегетативной нервной системы,
III- глазодвигательный нерв; YII - лицевой нерв; IX - языкоглоточный нерв; X - блуждающий нерв. 1 - ресничный узел; 2 - крылонебный узел; 3 - ушной узел; 4 - подниж-нечелюстной узел; 5 - подъязычный узел; 6 - парасимпатическое крестцовое ядро; 7 - экстрамуральный тазовый узел. Рис. 44. Парасимпатическая часть вегетативной нервной системы.
Автономная нервная система включает отделы как центральной, так и периферической нервной системы. В отличие от соматической, в автономной нервной системе эфферентная часть состоит из двух нейронов: преганглионарного и постганглионарного. Преганглионарные нейроны расположены в центральной нервной системе. Постганглионарные нейроны участвуют в образовании автономных ганглиев. В автономной нервной системе различают симпатический и парасимпатический отделы. В симпатическом отделе преганглионарные нейроны находятся в боковых рогах спинного мозга. Аксоны этих клеток (преганглионарные волокна) подходят к симпатическим ганглиям нервной системы, расположенным по обе стороны позвоночника в виде симпатической нервной цепочки. В симпатических ганглиях располагаются постганглионарные нейроны. Их аксоны выходят в составе спинномозговых нервов и образуют синапсы на гладких мышцах внутренних органов, желез, стенок сосудов, кожи и других органов. В парасимпатической нервной системе преганглионарные нейроны располагаются в ядрах ствола мозга. Аксоны преганглионарных нейронов идут в составе глазодвигательного, лицевого, языкоглоточного и блуждающего нервов. Кроме того, преганглионарные нейроны находятся также в крестцовом отделе спинного мозга. Их аксоны идут к прямой кишке, мочевому пузырю, к стенкам сосудов, снабжающих кровью органы, расположенные в области таза. Преганглионарные волокна образуют синапсы на постганглионарных нейронах парасимпатических ганглиев, расположенных вблизи эффектора или внутри него (в последнем случае парасимпатический ганглий называют интрамуральным). Все отделы автономной нервной системы подчинены высшим отделам центральной нервной системы. Отмечен функциональный антагонизм симпатической и парасимпатической нервной системы, что имеет большое приспособительное значение (см. табл. 1).
|
||||||
|
|
Последнее изменение этой страницы: 2019-12-25; просмотров: 323; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.16 (0.011 с.) |