Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Цельнометаллическая оболочкаСодержание книги
Поиск на нашем сайте
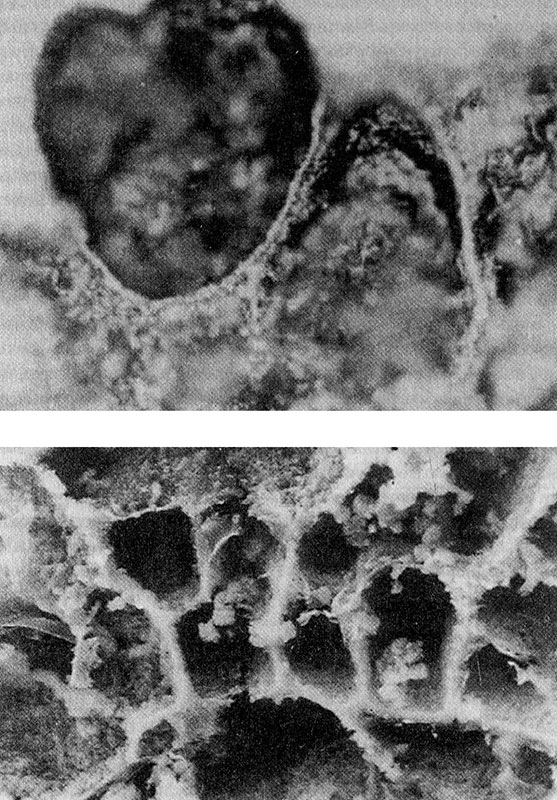
Мысль о том, что железо-серные минералы, такие как пирит (серный, или железный колчедан), играли какую-то роль в происхождении жизни, восходит к концу 1970-х гг., когда на океанском дне на глубине трех километров были открыты «черные курильщики». Эти серные гидротермальные источники представляют собой огромные, шаткие черные башни, перегретые при высоких давлениях морского дна и извергающие «черный дым» в окружающий океан. «Дым» состоит из вулканических газов и минералов, включая сернистое железо и сероводород, которые оседают в виде железосерных минералов. Самое удивительно в том, что черные курильщики, несмотря на высокую температуру и давление, а также абсолютную темноту, полны жизни. Там процветает целое экологическое сообщество, члены которого получают энергию непосредственно из гидротермальных источников и на первый взгляд независимы от Солнца[36]. Железо-серные минералы могут катализировать органические реакции, что они и делают по сей день в составе простетических групп многих ферментов, таких как железо-серные белки. Гипотеза о том, что железо-серные минералы были повитухами жизни, катализируя восстановление углекислого газа с образованием разнообразных органических молекул в адских условиях черных курильщиков, была разработана немецким химиком и по совместительству патентным поверенным Гюнтером Вэхстерхойзером в серии блестящих работ конца 1980-х и 1990-х гг. (Как заметил один ученый, при чтении этих работ возникает чувство, что они вывалились в наш мир через искривление во времени из конца XXI в.) Вэхстерхойзер предположил, что первые органические реакции происходили на поверхности железо-серных минералов. Сначала казалось, что его идеи согласуются с филогенетическими деревьями, построенными с использованием данных молекулярной генетики. Согласно этим деревьям, гипертермофилы (микробы, процветающие при высоком давлении и обжигающих температурах) принадлежат к числу наиболее древних групп как бактерий, так и архей. Впоследствии, однако, эти генетические данные стали казаться менее убедительными (см., например, книгу де Дюва), а реакции, о которых говорил Вэхстерхойзер, подверглись критике в плане термодинамики. Однако самое важное возражение против гипотезы черных курильщиков, возможно, связано с проблемой разбавления. Прореагировав на двухмерной поверхности кристалла, вещества отделяются от нее, и ничто не мешает им рассеяться по океанским просторам. Ничто не удерживает их вместе, если они не остаются прикрепленными к поверхности, а представить себе, что циклы биохимических реакций, всегда протекающих в растворах, эволюционировали в фиксированном положении на поверхности минерала, довольно трудно. В конце 1980-х гг. Майк Расселл предложил альтернативную серию идей; с тех пор он развивает их, в основном в сотрудничестве с Биллом Мартином. Расселла интересуют не столько огромные и страшные «черные курильщики», сколько более скромные места, где просачиваются на поверхность вулканические продукты. Одно из таких мест — отложения пиритов около деревни Тина (Ирландия), образовавшиеся 350 миллионов лет назад. Минералы образовали там огромное количество трубчатых структур, размером примерно с колпачок от ручки, а также пористые отложения, которые, по мнению Расселла, похожи на первые инкубаторы жизни. Наверное, говорит Расселл, эти пузырчатые структуры образовались при смешении двух химически несхожих жидкостей: горячих, восстановленных щелочных вод, просачивавшихся из глубин земной коры, и более окисленных, кислотных вод верхних слоев океана, содержавших углекислый газ и соли железа. Железо-серные минералы, такие как макинавит (FeS), оседали на микроскопических пористых мембранах в зоне смешивания. Это не просто спекуляции. Расселл и его давний соавтор Алан Холл воспроизвели этот процесс в лабораторных условиях. Впрыскивая раствор сульфида натрия (имитирующего просачивающуюся из земных глубин гидротермальную жидкость) в раствор хлорида железа (имитирующего древний океан) Расселл и Холл получили множество мелких, микроскопических пузырьков, окруженных железо-сульфидными мембранами (рис. 8).
Рис. 8. Древние клетки с железо-серными мембранами, (а) Электронная микрофотография тонкого среза железо-серного минерала (пирита) из окрестностей деревни Тина (Ирландия), образовавшегося 360 миллионов лет назад, (б) Электронная микрофотография структур, образовавшихся в лабораторных условиях после впрыскивания раствора сульфида натрия (NaS), имитирующего гидротермальную жидкость, в раствор хлорида железа (FeCl2), имитирующего богатый железом древний океан
У этих пузырьков есть две удивительные особенности, которые убеждают меня в том, что Расселл и Холл мыслят в правильном направлении. Во-первых, эти пузырьки по природе своей «хемиосмотичны»: снаружи среда более кислая, чем внутри. Это похоже на эксперимент Ягендорфа — Юрайба, в котором разницы pH по разные стороны мембран оказалось достаточно для производства АТФ. Поскольку эти пузырьки Расселла обладают естественным градиентом pH, то все, что нужно им для производства АТФ, — это включить в мембрану АТФазу. Это куда проще, чем выработать в процессе эволюции целый функциональный путь брожения! Если для того, чтобы сделать первый шаг к возникновению жизни, нужна была только АТФаза, то провидческие слова Рэкера о том, что АТФаза — это «элементарная частица жизни», оказались даже более справедливыми, чем он мог себе представить. Во-вторых, железо-серные кристаллы в мембранах пузырьков (как и железо-серные белки, которые по сей день входят в состав митохондриальных мембран) проводят электроны. Восстановленные жидкости, поднимающиеся из земной мантии, богаты электронами, а относительно окисленные океанские воды электронами бедны, и в результате возникает трансмембранная разность потенциалов, равная примерно нескольким сотням милливольт, что очень похоже на напряжение современных бактериальных мембран. Это напряжение стимулирует поток электронов через мембрану из одного компартмента в другой. Более того, поток отрицательно заряженных электронов притягивает положительно заряженные протоны изнутри, порождая зачаточный механизм закачивания протонов. Железо-серные «клетки» не только постоянно генерируют энергию, но и действуют как миниатюрные электрохимические реакторы, катализируя фундаментальные биохимические реакции и накапливая их продукты. Базовые строительные блоки жизни, включая РНК, АДФ, простые аминокислоты, маленькие пептиды и т. д., могли образоваться благодаря каталитическим свойствам железо-серных минералов, а также, возможно, осадочных глин, в тех самых реакциях, которые описал Гюнтер Вэхстерхойзер. Однако у гипотезы Расселла и Холла есть два огромных преимущества: участвующие в реакциях вещества и продукты реакций концентрируются около мембран, которые не дают им рассеиваться в океане, а сами реакции осуществляются за счет естественного источника энергии, протонного градиента.
Сама жизнь
Так ли все, о чем шла речь выше, сомнительно? В предыдущей главе я предположил, что происхождение жизни было куда более вероятным, чем происхождение эукариот. Подумайте о том, как это могло бы быть. Условия, подобные тем, что я только что описал, не были редкими на древней Земле. По оценкам ученых, вулканическая активность в те времена была в пятнадцать раз выше, чем сейчас. Земная кора была тоньше, океаны мелководнее, тектонические плиты только-только начинали формироваться. Продукты вулканической деятельности должны были просачиваться на поверхность во многих местах, не говоря уже о более активных вулканических процессах. Для образования миллионов мельчайших клеток, ограниченных железосерными мембранами, требовалась всего лишь разница окислительно-восстановительного потенциала и кислотности между океанской водой и вулканическими жидкостями, просачивавшимися из земных глубин, а такая разница, несомненно, существовала. Древнейшая Земля, какой представляет ее Расселл, была гигантской электрохимической батареей, окислявшей океаны за счет солнечной энергии. Ультрафиолетовые лучи расщепляли воду и окисляли железо. Образовавшийся из воды водород не удерживался земным притяжением и испарялся в космическое пространство. Океан постепенно окислялся, все больше отличаясь от преимущественно щелочной мантии. Согласно основным законам химии, в зонах смешивания неизбежно образовывались «клетки» со своим хемиосмотическим и окислительно-восстановительным градиентом. Смешиванию способствовала большая амплитуда колебаний уровня океана (приливно-отливного цикла), обусловленная силой тяготения недавно образовавшейся Луны, которая в то время была ближе к Земле, чем сейчас. Скорее всего, такие «клетки» действительно образовывались, возможно, массово. От них остались геологические свидетельства, такие как отложения в Тине. Отсюда далеко даже до первых бактерий, но для начала это совсем неплохо. Дело не только в том, что необходимые условия могли сложиться, но и в том, что они были устойчивы и существовали долгое время. Все, что было нужно, — это энергия Солнца; никаких тебе сомнительных нововведений вроде фотосинтеза или брожения. Нужно было, чтобы Солнце окисляло океаны, а оно именно это и делает. Ученые, бурно обсуждавшие различные формы энергии, возможные на древней Земле, — метеоритные удары, вулканическое тепло, молнии, — почему-то забывали про энергию Солнца (в отличие от доисторических мифов, которые всегда уделяли Солнцу ключевую роль). Как писал выдающийся микробиолог Фрэнклин Харольд в классической книге The Vital Force («Жизненная сила») (из почтения к нему я включил это выражение в название этой части книги): «Нельзя не прийти к мысли о том, что великий поток энергии, пронизывающий Землю, играет в биологии более важную роль, чем известно нашим современным мудрецам: возможно, что этот поток энергии не только позволил жизни возникнуть, но и непосредственно создал ее». Сотни миллионов лет Солнце служило постоянным источником энергии, необходимой для «расплаты» со вторым законом термодинамики. Оно создало химическое неравновесие и способствовало образованию естественных хемиосмотических «клеток». Эти изначальные условия до сих пор отражены в почти неизменном виде в фундаментальных свойствах всех современных клеток. Как органические клетки, так и их неорганические прототипы окружены мембраной, которая физически отграничивает органические компоненты клетки, не давая им раствориться в окружающем океане. В обоих случаях биохимические реакции катализируются минералами (в органических клетках они включены в состав простетических групп ферментов). В обоих случаях мембрана служит как барьером, так и переносчиком энергии, а энергия запасается в форме хемиосмотического градиента: снаружи есть положительный заряд и кислая среда, а внутри — относительно отрицательный заряд и щелочные условия. В обоих случаях окислительно-восстановительные реакции, транспорт электронов и закачка протонов восстанавливают этот градиент. Когда бактерии и археи наконец покинули инкубатор и пустились в путь по открытому океану, они унесли с собой нестираемую печать своего происхождения. Они гордо несут ее и поныне. Однако эта печать, напоминающая о происхождении жизни, также наложила на нее непреодолимое ограничение. Почему бактерии не вышли за рамки бактериального уровня организации? Почему за четыре миллиарда лет не возникла ни одна истинно многоклеточная, разумная бактерия? Если конкретнее: почему для возникновения эукариот понадобился союз архея и бактерии, а не просто постепенное усложнение какой-нибудь особо удачной эволюционной линии бактерий или архей? В третьей части книги мы увидим, что ответ на эту старинную загадку и объяснение удивительного расцвета эукариотической линии, давшей миру растения и животных, следует искать в самой природе производства энергии за счет создания трансмембранного хемиосмотического градиента.
Часть 3 Инсайдерская сделка [37] Основы сложности
Бактерии царили на Земле два миллиарда лет. В процессе эволюции они достигли почти безграничного биохимического разнообразия, но так и не приобщились к тайне большого размера и морфологической сложности. Возможно, эволюция жизни на других планетах остановилась на этом же уровне. Здесь, на Земле, большие размеры тела и высокий уровень сложности стали возможны только после того, как процесс производства энергии переместился в митохондрии. Что же помешало бактериям делегировать производство энергии каким-нибудь внутренним структурам? Подсказкой послужит нам один любопытный факт: митохондрии эукариот вот уже два миллиарда лет упорно сохраняют свою ДНК.
Клетка, полная «всяких штучек». Процесс производства энергии у эукариот приурочен к митохондриям
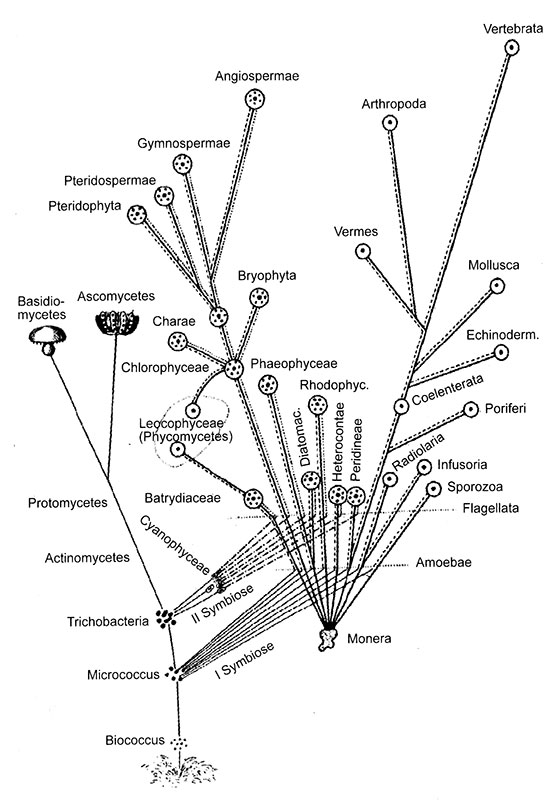
Вот список слов, при одном упоминании которых поперхнется пивом любой эволюционный биолог: целесообразность, телеология, лестница восходящей сложности, недарвиновская эволюция. Все эти термины ассоциируются с религиозным подходом к эволюции — верой в то, что возникновение и дальнейшее усложнение жизни были предначертаны, а человечество занимает промежуточное положение между низшими животными и ангелами в великой «цепи бытия», восходящей к Творцу. Теперь среди сторонников этого подхода можно найти не только теологов, но и астробиологов. Действительно, отрадно думать, что физические законы Вселенной благоприятствуют возникновению жизни, а отсюда недалеко до мысли, что человеческое сознание было почти неизбежным следствием этих же законов. Я уже оспаривал это положение в начале книги, и мы снова вернемся к этой теме в третьей части, где идет речь о происхождении биологической сложности. В первой части книги мы видели, что все сложные многоклеточные организмы состоят из эукариотических клеток, а бактерии за четыре миллиарда лет остались клетками-одиночками. Между бактериальными и эукариотическими клетками лежит огромная пропасть, и не исключено, что в других местах Вселенной жизнь не поднялась выше бактериального уровня организации. Мы видели, что эукариотическая клетка была плодом маловероятного союза бактерии и архея. Давайте теперь поищем первые ростки сложности. Какие именно особенности эукариотической клетки способствовали усложнению? Пусть это впечатление и обманчиво, но при взгляде на великое полотно эволюции после появления эукариотической клетки действительно возникает ощущение целенаправленности. Образ стремящейся к Богу цепи бытия, даже если он ложен, появился не на ровном месте. В этой части книги мы увидим, что семена сложности заронили в эукариотический мир митохондрии, ведь неуклонное усложнение жизни началось именно после их появления. Сложность не была предписана сверху. Она прорастала изнутри. В знаменитой книге Chance and Necessity («Случайность и необходимость») Жак Моно — молекулярный биолог, убежденный атеист и лауреат Нобелевской премии[38]— обсуждает тему целесообразности. Бессмысленно говорить о сердце, пишет он, умалчивая о том, что это насос, функция которого — обеспечивать ток крови по кровеносным сосудам. Но говоря так, мы, по сути, имеем в виду, что сердце служит определенной цели. А если мы скажем, что сердце возникло для того, чтобы обеспечивать ток крови, то окончательно впадем в телеологическую ересь, ведь тем самым мы припишем эволюционной траектории конечную цель. Тем не менее сердце вряд ли возникло для чего-то другого; если оно возникло не для того, чтобы обеспечивать кровоток, то просто чудо, что оно так прекрасно приспособлено для этой цели. Моно утверждал, что биология полна целесообразности и целенаправленности, и отрицать это было бы просто глупо, наша задача — это объяснить. Для этого мы должны ответить на следующий вопрос: как слепой случай — случайный механизм, не обладающий даром предвидения, — порождает совершенные и целесообразные биологические машины? Ответ на этот вопрос дал Дарвин, и он гласит, как мы знаем: «естественный отбор». Слепой случай всего лишь создает случайную изменчивость в популяции. Отбор не слеп или, по крайней мере, не случаен: он отбирает организмы в соответствии с их общей приспособленностью к определенной окружающей среде («выживание наиболее приспособленных»). Такие организмы передают потомству свою успешную генетическую организацию. Любые изменения, которые улучшают способность сердца обеспечивать ток крови, будут переданы следующим поколениям, а любые изменения, которые этому препятствуют, будут отброшены. В каждом поколении (в природных условиях) выживают и успешно размножаются только несколько процентов особей в популяции, и это, как правило, самые удачливые или самые приспособленные ее члены. На протяжении многих поколений фактор удачливости компенсируется, так что естественный отбор оставляет наиболее приспособленных из наиболее приспособленных, неизбежно совершенствуя конкретную функцию до тех пор, пока другие факторы отбора не начнут действовать в другом направлении. Таким образом, естественный отбор действует как храповик, придающий направленность случайной изменчивости. То, что получается в результате, действительно похоже на лестницу восходящей сложности. По большому счету биологическая приспособленность записана в генах, потому что только они передаются следующему поколению (ну, строго говоря, не только — еще передаются митохондрии). Действие естественного отбора на изменения генетической последовательности из поколения в поколение приводит к мелким усовершенствованиям, а в результате мы созерцаем головокружительный готический собор биологической сложности. Дарвин ничего не знал о генах, но с открытием генетического кода тут же стал понятен механизм возникновения случайной изменчивости в популяции: изменения последовательности «букв» ДНК за счет мутаций могут менять последовательность аминокислот в белках, что может положительно или отрицательно сказываться на их функции (или никак не сказываться на ней). Подобная изменчивость может возникать за счет одних только ошибок при копировании генов. В каждом поколении возникает порядка нескольких сотен мелких изменений последовательности ДНК (а она состоит из нескольких миллиардов «букв»), которые могут влиять, а могут не влиять на приспособленность. Мелкие изменения действительно происходят, подкидывая сырье в топку медленного эволюционного процесса, каким представлял его себе Дарвин. Наблюдать этот процесс в действии мы можем, проанализировав постепенные расхождения генетических последовательностей разных видов на протяжении сотен миллионов лет. Однако мелкие мутации — не единственный путь изменения генома (полной библиотеки генов организма), и чем дальше продвигается геномика (наука, изучающая геномы), тем менее важным представляется их значение. Для усложнения нужно, как минимум, больше генов — маленький бактериальный геном не мог бы кодировать человеческий организм и уж тем более не мог бы обеспечить бесчисленные генетические различия между разными людьми. В общем, при изучении разных видов просматривается корреляция между степенью сложности и числом генов (но не общим количеством ДНК). Откуда же берутся все эти «дополнительные» гены? Они возникают за счет удвоения существующих генов (или даже целых геномов), или за счет объединения двух и более разных геномов, или за счет распространения повторяющихся последовательностей ДНК (эти «эгоистичные» на первый взгляд последовательности, многочисленные копии которых разбросаны по геному, впоследствии могут пригодиться для чего-нибудь полезного — я имею в виду полезного для организма в целом). Ни один из этих процессов не является «дарвиновским» в строгом смысле слова, то есть не приводит к мелким постепенным улучшениям существующего генома. Речь идет о крупномасштабных изменениях количества ДНК — гигантских скачках через генетическое пространство, одномоментных кардинальных трансформациях генетических последовательностей (даже если они приводят к появлению не самих новых генов, а материала для них). Однако если отвлечься от масштаба изменений, в остальном этот процесс вполне согласуется с теорией Дарвина. Такие изменения генома по сути случайны, и в дальнейшем на них начинает действовать естественный отбор. Новые последовательности генов подстраиваются к выполнению новых задач за счет мелких изменений, накапливающихся из поколения в поколение. Короче говоря, если такие масштабные изменения количества ДНК не порождают нежизнеспособных монстров, они вполне имеют право на существование. Если окажется, что от удвоения количества ДНК никакого особого проку нет, естественный отбор наверняка избавится от излишков. Чем больше «ненужной» ДНК будет «выброшено за борт», тем ниже окажется потолок потенциальной сложности организма, так как при этом пропадает материал, необходимый для образования новых генов, а сложным организмам, как мы знаем, нужно много генов. Тут мы снова возвращаемся к лестнице восходящей сложности. Вспомним, как непохожи эукариоты и бактерии. Бактерии за четыре миллиарда лет остались бактериями: достигнув исключительного разнообразия и сложности в биохимическом плане, они не произвели решительно никакой истинной морфологический сложности. Если смотреть только на размер, форму тела и внешний вид, то можно сказать, что они вообще не изменились. Эукариоты же очень существенно усложнились за половину отведенного бактериям срока. У них возникли внутренние мембранные системы, специализированные органеллы, хитроумные клеточные циклы, заменившие простое деление клеток, пол, огромные геномы, фагоцитоз, хищничество, многоклеточность, дифференциация, большие размеры тела и, наконец, такие потрясающие инженерные решения, как полет, зрение, слух, эхолокация, мозг и сознание. Поскольку эти усложнения появлялись с течением времени, их вполне можно расположить в виде лестницы восходящей сложности. Итак, мы видим бактерии, с их почти бесконечным биохимическим разнообразием, но полным отсутствием тенденции к усложнению, и эукариот, довольно однообразных в биохимическом плане, но восхитительно непохожих друг на друга в том, что касается морфологии. Убежденный дарвинист, которому указали бы на пропасть между бактериями и эукариотами, мог бы ответить: «Ну что вы, бактерии все-таки усложнились — разве не от них произошли первые эукариоты, которые, в свою очередь, дали начало всем тем организмам, исключительную сложность которых вы превозносите?» Это справедливо, но только в некотором смысле, и в этом-то вся загвоздка. Митохондрии, как я покажу чуть ниже, могли возникнуть только за счет эндосимбиоза — объединения двух геномов в одной клетке, гигантского скачка через генетическое пространство, — а сложные эукариотические клетки вообще не могли возникнуть без митохондрий. Подобная точка зрения основана на представлениях о том, что эукариотическая клетка сформировалась в процессе союза, который дал начало и митохондриям тоже, и что обладание митохондриями является, или являлось в прошлом, необходимым условием эукариотической природы. Это представление расходится с традиционными взглядами на происхождение эукариот, поэтому я позволю себе в двух словах напомнить вам, почему это важно. В первой части книги мы рассмотрели происхождение эукариотической клетки согласно взглядам Тома Кавалье-Смита — именно они лучше всего отражают традиционную точку зрения. Давайте вкратце перечислим основные пункты: прокариотическая клетка утратила клеточную стенку (возможно, под влиянием выделяемых другими бактериями антибиотиков), но выжила, так как уже имела внутренний белковый скелет (цитоскелет). Утрата клеточной стенки коренным образом изменила образ жизни и способ размножения клетки. У нее появилось ядро и сложный жизненный цикл. За счет цитоскелета она смогла двигаться и менять форму тела, как амеба, и перешла к новому, хищному образу жизни, заглатывая крупные пищевые частицы, например целые бактерии, путем фагоцитоза. Короче говоря, первая эукариотическая клетка приобрела ядро и эукариотический образ жизни за счет обычной дарвиновской эволюции. На относительно поздней стадии одна такая клетка случайно заглотила пурпурную бактерию, возможно паразита вроде Rickettsia. Оказавшиеся внутри бактерии выжили и постепенно превратились, опять же за счет обычной дарвиновской эволюции, в митохондрии. Обратите внимание на две особенности этой теории: во-первых, ее явный «дарвиновский крен», так как факту союза двух разных геномов — по сути, недарвиновского способа эволюции — уделяется мало внимания; а во-вторых, то, что она умаляет значение митохондрий. Согласно этой теории, митохондрии «подключились» к полноценной эукариотической клетке и были утрачены во многих примитивных линиях (например, у Giardia). Они являются эффективным способом производства энергии, и не более того. Просто новой клетке взамен устаревшего моторчика поставили двигатель от «порше». По-моему, такой подход крайне плохо объясняет, почему все сложные клетки имеют митохондрии или, если посмотреть с другой стороны, почему митохондрии необходимы для эволюции сложности. Теперь обратимся к водородной гипотезе Билла Мартина и Миклоша Мюллера, которую мы тоже обсуждали в первой части. Согласно этой радикальной гипотезе, тесные взаимоотношения между двумя очень разными прокариотическими клетками были изначально основаны на химической взаимозависимости. В конце концов одна клетка физически захватила другую, и в ней оказались два генома. Этот гигантский скачок через генетическое пространство породил многообещающего монстра, и он тут же подпал под давление естественного отбора, которое привело к переносу генов «гостя» к хозяину. Принципиальный момент водородной гипотезы заключается в том, что примитивного эукариотического организма, который обладал ядром и вел хищный образ жизни, но не имел митохондрий, — никогда не было. Первый эукариот родился от союза двух прокариот, и это было абсолютно недарвиновским явлением. Маршрут был преодолен за один бросок без перевалочных пунктов. Все это ставит с ног на голову наши обычные представления об эволюции как о ветвящемся древе жизни. В этом можно убедиться, посмотрев на древо жизни, изображенное русским биологом Константином Мережковским в 1905 г. (рис. 9).
Рис. 9. Предложенное Мережковским инвертированное древо жизни. Обычное «дарвиновское» древо жизни всегда строго дихотомично: ветви ветвятся, но никогда не сливаются. Эукариотическая клетка произошла за счет эндосимбиоза. На древе жизни это показано обратной бифуркацией: ветви сливаются, отчасти инвертируя древо жизни
Древа жизни всегда вызывали много споров; особенно яростно их критиковал Стивен Джей Гулд, утверждавший, что после открытия кембрийского взрыва о традиционном древе жизни говорить просто нельзя. Кембрийский взрыв — это великое и по геологическим меркам очень быстрое увеличение биоразнообразия примерно 560 миллионов лет назад. Впоследствии большинство крупных ветвей были безжалостно обрезаны — целые типы животных вымирали без следа. Дэниел Деннет в своей книге «Опасная идея Дарвина» разносит в пух и прах якобы радикальные идеи Гулда, утверждая, что его вариант не отличается от прежних ничем, кроме деформации осей: вместо горделиво возвышающегося дерева мы имеем дело с низкорослым кустарничком, пустившим вверх несколько чахлых побегов. Но Мережковскому подобные упреки не грозят. Его эволюционное древо действительно перевернуто — новый домен жизни возникает за счет срастания, а не ветвления. То, о чем я сейчас говорю, не является откровением. Все эти соображения на слуху, а симбиоз входит в традиционный эволюционный канон, даже если там ему отводится всего лишь роль механизма возникновения эволюционных новшеств. Например, Джон Мейнард Смит (великий ученый, к сожалению, недавно умерший) и Эрш Сатмари в интереснейшей книге The Origins of Life («Истоки жизни») сравнивают биологический симбиоз с мотоциклом — симбиозом велосипеда и двигателя внутреннего сгорания. Даже если считать симбиоз шагом вперед, довольно неуклюже шутят они, кто-то должен был сначала изобрести и велосипед и двигатель внутреннего сгорания. Так и в жизни: сначала естественный отбор создает части, а потом симбиоз творчески использует их. Итак, симбиоз лучше всего объясняется в терминах дарвиновской эволюции. Все это справедливо, но затеняет тот факт, что некоторые коренные эволюционные новшества были возможны только за счет симбиоза. Продолжая метафору Мейнарда Смита и Сатмари, если велосипед и двигатель внутреннего сгорания могли возникнуть независимо путем естественного отбора, то также мог бы возникнуть и сам мотоцикл. Конечно, при наличии готовых составных частей мотоцикл появился бы гораздо быстрее, но, если времени достаточно, он вполне мог бы возникнуть сам по себе без помощи симбиоза. Я считаю, что эукариотическая клетка сама по себе возникнуть не могла. Предоставленные самим себе, бактерии не могли дать начало эукариотам за счет одного только естественного отбора: симбиоз был нужен для преодоления пропасти между бактериями и эукариотами, а митохондрии были необходимы для разбрасывания семян сложности. Сложная жизнь невозможна без митохондрий, а митохондрии — без симбиоза. Без митохондриального союза не было бы ничего, кроме бактерий. Независимо от того, считаем ли мы симбиоз дарвиновским процессом или нет, понимание того, почему симбиоз с митохондриями был необходим, равнозначно пониманию нашего прошлого и нашего места во Вселенной[39]. В третьей части книги мы увидим, почему между прокариотами и эукариотами лежит зияющая пропасть и почему только симбиоз мог перекинуть через нее мост. Учитывая механизм производства хемиосмотической энергии (см. вторую часть книги), происхождение эукариот от прокариот путем естественного отбора практически невозможно. Поэтому бактерии остались бактериями, и по этой же причине жизнь, какой мы ее знаем, то есть основанная на клетках, химии углерода и хемиосмосе, вряд ли преодолеет бактериальный уровень сложности где бы то ни было еще во Вселенной. Также в третьей части мы увидим, почему митохондрии заронили семена сложности на эукариотическую почву и поместили эукариот на стартовую позицию в основании лестницы восходящей сложности; а в четвертой части мы поймем, почему митохондрии обеспечили подъем эукариот по этой лестнице.
|
||
|
|
Последнее изменение этой страницы: 2019-04-27; просмотров: 220; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.115 (0.018 с.) |