
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Робота при перенесенні зарядів через мембрануСодержание книги Поиск на нашем сайте
Величина осмотичного тиску Розглянемо, який тиск здійснюють на мембрану з напівпроникненістю молекули розчинника і розчиненої речовини.
На рис. 8.1.2. дано пояснення походження осмотичного тиску. Молекули розчинника (маленькі сірі кружечки) вільно проходять крізь мембрану й тиску на неї не створюють (1). Молекули розчиненої речовини (великі порожні кружечки) не проходять крізь мембрану й здійснюють на неї такий самий тиск, який здійснював би газ на стінку посуду при такій самій концентрації цих молекул і температурі. З газових законів відомо, що цей тиск можна описати рівнянням:
де p - тиск, Па; V – об’єм, м3; m - кількість речовини, моль; R - газова постійна, Дж.моль-1.К-1; T - температура, K. У нашому випадку p - це осмотичний тиск. Величина
Примітка: молярна концентрація речовини зовсім не обов'язково дорівнює молярній концентрації осмотично-активних часток (яку називають ще осмотичною концентрацією або осмомолярністю). Солі, такі як NaCl або KCl у водному розчині повністю дисоціюють, і їх осмотична концентрація у два рази перевищує молярну концентрацію. Молекула CaCl2 дисоціює у водяному розчині на три части, тому її осмотична концентрація буде перевищувати молярну у три рази.
Види роботи:Механічна, розширення газу, осмотична, електрична. Осмотична робота й енергія Осмотична робота - це робота, яку треба зробити, щоб збільшити концентрацію речовини в даному розчині. Таку роботу буде робити гіпотетичний пристрій, зображений на 8.2.1. у третьому стовпчику. Він складається з циліндру, в якому роль поршня виконує пластинка з "напівпроникної речовини". Крізь пластинку може вільно проходити розчинник, але не можуть проходити молекули (іони) розчиненої сполуки. Приклавши до такої пластинки тиск p, можна стиснути об'єм розчину з вихідного V1 до кінцевого V2; робота, яка буде зроблена, як й у випадку стиску газу, дорівнюватиме:
12.
13. §5. 5. Анатомо-фізіологічні фактори, що визначають механічні властивості м’язів Як ми зазначали, рухова діяльність людини визначається, в основному, силою тяги збуджених м’язів, швидкістю і величиною їхнього скорочення. Довжина, сила і швидкість скорочення є найбільш важливими характеристиками механічних властивостей м’язів. Анатомо-фізіологічні фактори, що визначають силу м’язів. Сила тяги м’язів залежить від сукупності механічних, анатомічних і фізіологічних умов. Основною механічною умовою, що визначає тягу м‘язу, є навантаження. Без навантаження, зрозуміло, для м’язу не може бути його напруга, а через це – і сила тяги. Першим правилом м’язової механіки є принцип Вебера: Сила м’язів, при інших однакових умовах пропорційна його поперечному перетину. Суть цього правила полягає в тому, що чим більша кількість волокон входять до складу м’язу, тим більшою є його сила. Тобто, товсті волокна розвивають більш високу напругу, ніж тонкі. Одинока нейромоторна одиниця, яка включає, наприклад, 100 м’язових волокон, може розвити силу в 10-20 г. Багато скелетних м’язів володіють силою, яка переважає вагу тіла. Така велика сила необхідна в зв’язку з тим, що ланки рухового апарату у більшості випадків працюють за принципом важелів III роду, де шляхом програшу у силі досягається виграш у величині амплітуди руху і швидкості руху. Для характеристики здібності до напруги важливим є визначення абсолютної сили м’язу – це сила, що приходиться на 1 см2 поперечного перетину через м’язові волокна. Щоб її визначити, потрібно величину сили, що розвивається м’язом в цілому, поділити на величину її фізіологічного поперечника. Фізіологічний поперечник – площа поперечного перетину через всі волокна м’язу – не завжди співпадає з анатомічним поперечником (площа поперечного перетину через м‘яз). Співпадання спостерігається лише у паралельно-волокнистих м’язів. Для поперечно-волокнистих м’язів, по типу яких побудовано більшість скелетних м’язів, фізіологічний поперечник дуже перевищує анатомічний. Завдяки цьому ці м’язи є відносно більш сильними, ніж паралельно-волокнисті. Абсолютна сила м’язів людини має такі величини: для ікроножного – 6,24 кг, для розгиначів шиї – 9,0 кг, жувального – 10 кг, двоголового плеча – 11,4 кг, плечового – 12,1 кг, трьохголового – 16,87 кг. Це визначає анатомічні умови прояву сили тяги м’язів: фізіологічний поперечник визначає сумарну тягу всіх волокон з врахуванням їх взаємного розташування, тобто їх пружної деформації. Розташування м’язу у кожний момент руху визначає кут його тяги відносно кісткового важелю, тобто впливає на значення моменту сили тяги м’язу. При кутах, що відрізняються від прямого, крім обертальної, є ще і підкріплююча складова тяги м’язу. З появою підкріплюючої складової зменшується обертальна. Фізіологічні умови прояву тяги м’язу, в основному можна звести до його збудження і стомлення, які відображуються на можливостях м’язів чи підвищуючи, чи знижуючи його напругу. Крім того, сила напруги м’язу може бути різною в залежності: · від впливу на неї нервових імпульсів: центральна нервова система може парирувати силу напруги м’язу як через залучення в роботу різної кількості рухових (моторних) одиниць, так і через зміну частоти імпульсів, що вона посилає. Тобто сила тяги м’язового скорочення залежить від кількості моторних одиниць, які залучаються одночасно і від частоти збудження кожної з них; · від вихідної довжини м’язу: якщо попередньо зблизити кінці м’язу, то він, при інших однакових умовах, буде розвивати меншу напругу. І, навпаки, якщо м’яз попередньо розтягнути, то він становиться здібним розвинути більш високу напругу. Однак при дуже великому розтягуванні м’язова сила знов падає. У стані спокою м’язи не є повністю розслабленими, а зберігають деяку напругу, яка називається тонусом (степінь пружності м’язу). Анатомо-фізіологічні фактори, що визначають швидкість скорочення м’язів. Величина тяги м’язу пов’язана зі швидкістю його деформації. Це залежить: · від величини внутрішнього тертя, тобто в’язкості. М’язи, що складаються зі “швидких” волокон, наприклад, ікроножний, скорочується з більш великою швидкістю, ніж ті, що побудовані з “повільних” волокон. Швидкі і повільні волокна згруповані у різні нейромоторні одиниці. Нервові центри, приводячи у рух то швидкі, то повільні одиниці, можуть парирувати швидкість скорочення. · Від сили, яка розвивається м’язами · Від навантаження. Чим більша сила напруження і чим менша маса частин тіла, що переміщується, тим більша швидкість скорочення (при інших однакових умовах). Ця закономірність визначається кривою Хілла (рис.4.4.1.2.). При переборювальній його роботі при підвищенні швидкості скорочення м’язу його гранична напруга спадає. При роботі по виконанню відступальних дій підвищення швидкості розтяжіння м’язу підвищує його напругу. Це, як вказао вище, дуже важливо при оцінці напруги м’язу при швидких рухах. Анатомо-фізіологічні фактори, що визначають величину скорочення м’язу. Тут діє принцип Бернуллі: Величина скорочення м’язу пропорційна довжині його волокон. Найбільше за величиною скорочення дають м’язи, які побудовані з довгих волокон – паралельно-волокнисті. Скорочення м’язу залежить також від сили, що виникає у ньому при збудженні, і від величини вантажу, який він переміщує. Чим більшою є активна сила м’язу і чим меншою є протидіюча йому зовнішня сила, тим більшим є його скорочення. Сила напруги, величина і швидкість скорочення м’язу не є константами. У початковий період м’язової роботи відбувається підвищення сили напруги, швидкості і величини скорочення м’язів. При стомленні ці показники зменшуються. Також вони зменшуються через систематичні заняття фізичними вправами.
Режими роботи м’язів Розрізняють 3 основних види таких режимів: - ізометричний; - ізотонічний (анізометричний); - ауксотонічний. При ізометричному (підтримуючому) режимі довжина м’язу не змінюється (від грецького «ізо» - однаковий, «метр» - довжина). При ізотонічному скороченні м’яз скорочується до деякої довжини, а потім розслаблюється, подовжується. Наприклад, у режимі ізометричного скорочення працюють м’язи людини, яка підтягнулася і утримує своє тіло у цьому положенні; або при утриманні штанги; жувальні м’язи при стиснутих щелепах, скорочення міокарду шлуночків при зачинених клапанах – це також приклад ізометричного режиму роботи м’язу. У цьому режимі функціонують м’язи бігуна, плавця, велосипедиста і.т.ін.(скорочення біцепсу плеча). При цьому скороченні кут тяги м’язу змінюється => змінюється момент сили м’язу і момент сили опору (навантаження). У цих умовах зберегти напругу м’язів постійною дуже важко, та і необхідності у цьому теж немає. При цьому режимі маємо наступне: чим більшим є навантаження F, тим меншим є скорочення м’язу і коротшим є час, наприклад, утримання якогось вантажу при його підйомі. При деякому навантаженні F=Fо м’яз зовсім перестає піднімати об’єкт. Це значення є максимальною силою ізометричного скорочення для певного м’язу. На кривій Хілла (рис.5.4.1.1.) ізометричному режиму відповідає величина статичної сили (F0), при якій скорочення м’язу дорівнює 0.
Рис.5.4.1.1. Взаємозв’язок між силою F і швидкістю скорочення V м’язу (крива Хілла)
У рухах людини звичайно має зміна і довжини, і напруги м’язів. Це ауксотонічний режим. У цьому випадку довжина м’яза і його напруга змінюються під впливом багатьох факторів (як біологічних, так і механічних). У анізометричного (ізотонічного) режиму 2 види. У переборювальному (переможному) режимі довжина м’язу зменшується у результаті скорочення, а в уступальному режимі м’яз розтягується зовнішньою силою. Наприклад, ікроножний м’яз функціонує у режимі уступання при взаємодії ноги з опорою у фазі амортизації, а у режимі перемоги – у фазі відштовхування. Права частини кривої Хілла (див. рис. 5.4.1.1.) відображує закономірності переборювальної роботи, при якій зростання швидкості скорочення м’язу викликає зменшення сили тяги. У режимі уступання спостерігається зворотня картина: підвищення швидкості розтягання м’язу супроводжується підвищенням сили тяги. Це є причиною чисельних травм у атлетів (наприклад, розрив ахілового сухожилля у спринтерів і стрибунів у довжину). Рівняння Хілла пов’язує між собою силу (F) і швидкість скорочення м’язу (v) при ізотонічному скороченні м’язу:
де: v – швидкість скорочення м’язу, F – м’язова сила або зовнішня сила прикладеного до нього навантаження, v max – максимальна швидкість скорочення м’язу, а - const, що мають розмірність сили, b - const, що має розмірність швидкості, Fо – максимальна сила ізометричного скорочення. 20. Розподіл тиску крові в судинній системі з деяким наближенням описується формулою Пуазейля:
де Q – об’ємна швидкість течії крові, r – радіус зазору судини, η – динамічна в’язкість крові (в Пуазах), l – довжина ділянки судини, ∆Р – перепад тисків на цій ділянці, R – гідравлічний тиск Звідси:
Тобто: 1. Об’ємна швидкість є прямо пропорційною зміні тисків і четвертій степені радіусу судини (r4)): так, при збільшенні радіусу на 16%, об’ємна швидкість течії рідини зростає на 100%. Тому незначні зміни ширини зазору кровоносних судини сильно позначаються на течії крові. Саме цьому природні механізми нервової і гуморальної регуляції кров’яного тиску, а також дія лікарських препаратів, які нормалізують тиск, спрямовані на зміну зазору судин. 2. Об’ємна швидкість є зворотно пропорційною довжині судини (l) і в’язкості рідини (η), 3. Гідродинамічний опір є прямо пропорційним довжині судини і в’язкості рідини, яка, в свою чергу, залежить від кількості еритроцитів, вмісту білка в плазмі та інших факторів. Гідродинамічний опір кровоносної судини є більшим, ніж у води. Периферійний опір судинної мережі людини дорівнює 1700 дин на с/см2 Різних тканин організму
При лікувальних процедурах електроди з прокладками накладаються в певних місцях на поверхню тіла. Електропровідність окремих ділянок організму, які знаходяться між електродами, значною мірою обумовлена електропровідністю шару шкіри і підшкіряно-жирової клітчатки, яка знаходиться безпосередньо під електродами, бо струм, який проходить крізь цій шар, розгалужується і проходить крізь більш заглиблені шари тканин багатьми паралельними гілками з найменшим опором. Тобто всередині організму струм розподіляється, здебільшого, через кровоносні і лімфатичні судини, м’язи, оболонки нервових стволів. Тому розгалуження струму в тканинах організму можуть бути складними і, навіть, захоплювати та райони, які далеко лежать від місця накладання електродів. Електропровідність шкіри, крізь яку струм проходить головним чином за каналами потових і сальних залоз, залежить від товщини і стану її поверхневого шару. Тобто і опір шкіри, в свою чергу, визначається її станом: товщиною, віком, вологістю та іншим (табл. 11.2.2.). Через це, електропровідність тканин і органів залежить від їх функціонального стану і тому може бути використана в якості діагностичного показника. Так, наприклад, це відбувається при запаленні, коли клітини набухають, через що зменшується перетин міжклітинних з’єднань і підвищується електричний опір; фізіологічні явища, які викликають пітливість, супроводжується зростанням електропровідності шкіри і, навпаки, суха загрубіла шкіра є поганим провідником т.ін.. Таблиця 11.2.2. Значення питомого опору різних тканин і рідин організму
Це свідчить, що моделювати електричні властивості біологічних тканин можна використовуючи резистори, які володіють активним опором, і конденсатори – носії ємкісного опору. В якості моделі, зазвичай, використовують еквівалентну електричну схему тканин організму. Це – схема, яка складається з резисторів і конденсаторів, частотна залежність (дисперсія) імпедансу[5] якої близька до частотної залежності імпедансу біологічної тканини. Таким чином, еквівалентна електрична схема ділянки тканин організму, що перебувають між накладеними на поверхню тіла електродами, повинна містити як електропровідні, так й ємнісні елементи, тобто вона може бути складена з резисторів і конденсаторів, включених між собою послідовно або паралельно. Найпоширенішими елементами в такій схемі є конденсатор С и резистор R', включені послідовно й спільно шунтировані резистором R (наприклад, для шару шкіри й підшкірної клітковини разом з контактним шаром між шкірою й електродом) (рис. 11.2.2, а), або конденсатор С и резистор R, включені паралельно (рис. 11.2.2, б). Загальний опір і загальна ємність ділянки тканин організму, що перебувають між електродами, у значній мірі залежать від їхньої площі (площа прокладок під електродами), щільності прилягання до поверхні тіла, ступеня зволоження прокладок і т.д., а також стану самих тканин (ступінь кровонаповнення, наявність набряклості й т.п.). В умовах лікувальних процедур цей опір при постійному струмі має порядок 1000-5000 0м, ємність - трохи сотих часток мікрофаради (при «точкових» електродах опір збільшується до десятків тисяч ом й ємність знижується до тисячних часток мікрофаради).
27. Безперервний постійний струм напругою 60-80 В використовують як лікувальний метод фізіотерапії – гальванізація. Джерелом струму звичайно служить двохнапівперіодний випрямувач – апарат для гальванізації. Застосовують для цього електроди із листового свинцю або станіолю 0,3-0,5 мм. Через те, що продукти електролізу розчину солей, який є в тканинах, викликає прижигання, то між електродами і шкірою розміщують гідрофільні прокладки, які змочуються, наприклад, теплою водою. Дозують силу струму за показниками міліамперметру, при цьому обов’язково враховують допустиму щільність струму – 0,1 мА/см2. Постійний струм широко використовується також для введення через шкіру і слизисті оболонки лікарських препаратів. Цей метод отримав назву електрофорезу лікарських речовин. Під електроди на шкіру кладуть прокладки, які змочуються відповідним лікарським препаратом. Через катод вводять аніони (йод, гепарин, бром), а через анод – катіони (Nа, Ca, Mg, новокаїн). Препарат вводять з того полюсу, який заряд він має: аніони вводять з катоду, катіони – з аноду. При лікувальному електрофорезі між електродами утворюється складний ланцюг, який складається з розчинів лікарських речовин, якими змочуються прокладки, і розчинів електролітів, які входять до складу тканин організму. На рис.12.2.1. умовно наведений об’єкт (електропровідні тканини організму, які вміщують розчин хлористого натрію), на які накладені прокладки П з розчином хлористого кальцію і під від’ємним – йодистого калію (напрямок руху іонів показаний стрілками). На ділянках а – е мають місце поляризаційні явища – скопичення іонів по обидві боки тканинних перегородок, які погано проводять струм. Електрофорез має ряд переваг у порівнянні зі звичайними методами введення лікарських препаратів, бо дозволяє вводити їх безпосередньо у тканини, мінуючи шлунково-кишковий тракт і кров.
|











 Тканини організму складаються зі структурних елементів – клітин, які омиваються тканинною рідиною. Такий елемент представляє собою два середовища, які відносно добре проводить струм (рідина тканин і цитоплазма клітини) та які розділені шаром, який погано проводить струм (клітинною мембраною) (рис.11.2.1., а). Через це така система має електричну ємність. При проходженні крізь тканини постійного струму в таких елементах відбувається накопичення по обидві боки мембрани іонів різного знаку, тобто утворюється система, що подібна до зарядженого конденсатора.
Тканини організму складаються зі структурних елементів – клітин, які омиваються тканинною рідиною. Такий елемент представляє собою два середовища, які відносно добре проводить струм (рідина тканин і цитоплазма клітини) та які розділені шаром, який погано проводить струм (клітинною мембраною) (рис.11.2.1., а). Через це така система має електричну ємність. При проходженні крізь тканини постійного струму в таких елементах відбувається накопичення по обидві боки мембрани іонів різного знаку, тобто утворюється система, що подібна до зарядженого конденсатора. В тканинах зустрічаються і макроскопічні утворенні, які складаються з різних з’єднувальнотканних оболонок і перегородок, тобто які є поганими провідниками, по обидва боки яких знаходяться тканини з великою кількістю рідини – тобто добрі провідники. Все це придає тканинам організму ємкісні властивості.
В тканинах зустрічаються і макроскопічні утворенні, які складаються з різних з’єднувальнотканних оболонок і перегородок, тобто які є поганими провідниками, по обидва боки яких знаходяться тканини з великою кількістю рідини – тобто добрі провідники. Все це придає тканинам організму ємкісні властивості.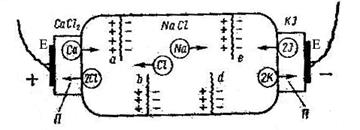
28. § 12.5. Механізм біологічної дії електромагнітних хвиль радіочастотного діапазону
Природними джерелами хвиль радіочастотного діапазону є Сонце, планети та інші космічні об’єкти. З них найбільшу інтенсивність має сонячне випромінювання, щільність потоку енергії якого при входженні до атмосфери Землі складає 1360 Вт/м2. У радіозв’язку, промисловості, медицині широко використовують штучні джерела радіовипромінювання. Вплив мікрохвильового випромінювання на біооб’єкти вивчають, використовуючи в якості джерел електромагнітного випромінювання клістрони, магнетрони та ін.
З усього радіохвильового спектра (табл.12.1.) виділяються дециметровий, або ультрависокочастотний (УВЧ) діапазон (λ =1...0,01м, υ =3.108...3.109Гц), і сантиметровий, або зверхвисокочастотний (ЗВЧ) діапазон (λ =10-1...10-2м, υ =3.109...3.1010Гц).
Мірою інтенсивності I мікрохвильового випромінювання є щільність потоку енергії випромінювання (модуль вектора Пойтинга):
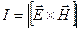
| (12.5.1.) |
де Е – напружність електричного поля, Н – напружність магнітного поля. Щільність потоку вимірюється у Вт/м2.
Іноді при дослідженні впливу випромінювання на речовину користуються питомими значеннями поглинутої одиницею об’єму або одиницею маси поглинаючої речовини потужності, що вимірюють відповідно у Вт/м3 або Вт/кг. Кількість поглинутої енергії одиницею маси об’єкта називають дозою опромінення (Дж/кг).
Електромагнітне випромінювання, особливо в ЗВЧ діапазоні, підвищує температуру тканини на деяку величину DT, причому DT є прямо пропорційною питомій поглиненій потужності P. Це дозволяє визначити поглинену потужність ЗВЧ-випромінювання в залежності від нагрівання тканини за наступною емпіричною формулою:
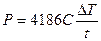
| (12.5.2.) |
де С – питома теплоємність речовини, t – час опромінення.
Енергія мікрохвильового випромінювання (1,24.10-6...1,24.10-4 еВ) є недостатньою для виникнення електронно-збуджених станів. Тому вплив мікрохвильового випромінювання на живі організми викликає теплові і нетеплові ефекти.
Тепловий ефект електромагнітного поля визначається його впливом як на іони, так і на нейтральні частинки. В першому випадку змінні поля викликають змінні струми провідності у тканинах, що супроводжується виділенням джоулева тепла. У другому випадку електричне поле викликає поляризацію нейтральних молекул і виникнення струмів зміщення.
Коливання частиць у змінному полі супроводжується тертям, а через це – виділенням теплової енергії. Зі зменшенням частоти зовнішнього поля відповідно зменшується частота коливань частиць і теплові втрати є меншими. З іншого боку, при збільшенні частоти електрони (у випадку деформаційної поляризації) або молекули (у випадку орієнтаційної поляризації) можуть не встигати змінювати своє положення у просторі, що також знижує теплові втрати. З цього маємо, що при деякому значенні частоти коливань тепловий ефект буде максимальним (резонанс поглинання). Це спостерігається у випадку співпадання частоти зовнішнього поля з зовнішньою частотою коливань частиці. Наприклад, у випадку орієнтаційної поляризації молекул води максимум поглинання знаходиться на частоті 1010Гц, тобто лежить в ЗВЧ діапазоні. Орієнтаційна поляризація молекул води вносить основний внесок у поглинання електромагнітного випромінювання живими організмами. Тому тканини, в яких вміст води є великим, нагріваються сильніше (наприклад, м’язи і кров) у порівнянні з тими, де вміст води є меншим (кісткова і жирова тканини).
Однією з важливих характеристик електромагнітного випромінювання є глибина його проникнення d у тканини. З підвищенням частоти випромінювання і діелектричної проникності тканини, глибина проникнення зменшується. Чим більшим є вміст води у тканині, тим меншим є d. Наприклад, глибина проникнення УВЧ-хвиль (υ =3.108...3.109Гц) в м’язовій тканині і шкіри складає біля 4 см, а в жировій і кістковій тканині – біля 20см. Ці ж показники для ЗВЧ-хвиль (υ =3.109...3.1010Гц) складають вже відповідно 2 і 10 см.
Нагрівання тканини традиційними способами і нагрівання, яке викликане впливом ЗВЧ-випромінювання, істотно різняться. В першому випадку відбувається нагрівання поверхневого шару, і проникнення тепла в більш глибокі шари залежить від теплопровідності тканини, в той час як УВЧ і ЗВЧ-опромінення викликає нагрівання всього об’єму опроміненого об’єкту. Зі зменшенням довжини хвилі зменшується глибина проникнення випромінювання у тканину, що викликає нагрівання лише поверхневих шарів.
Величина осмотичного тиску
Розглянемо, який тиск здійснюють на мембрану з напівпроникненістю молекули розчинника і розчиненої речовини.

| Рис. 8.1.2. Тиск, який здійснює на напівпроникну мембрану молекулами розчинника (маленькі сірі кружечки) і розчиненої речовини. Молекули розчинника вільно проходять крізь мембрану й тиски на неї не роблять (1). Молекули розчиненої речовини не проходять крізь мембрану й натискають на неї, рівне тиску газу на стінку. |
На рис. 8.1.2. дано пояснення походження осмотичного тиску. Молекули розчинника (маленькі сірі кружечки) вільно проходять крізь мембрану й тиску на неї не створюють (1). Молекули розчиненої речовини (великі порожні кружечки) не проходять крізь мембрану й здійснюють на неї такий самий тиск, який здійснював би газ на стінку посуду при такій самій концентрації цих молекул і температурі. З газових законів відомо, що цей тиск можна описати рівнянням:

| (8.1.1) |
де p - тиск, Па; V – об’єм, м3; m - кількість речовини, моль;
R - газова постійна, Дж.моль-1.К-1; T - температура, K.
У нашому випадку p - це осмотичний тиск. Величина  - це молярна концентрація осмотично-активних частинок, звідси знаходимо рівняння осмотичного тиску:
- це молярна концентрація осмотично-активних частинок, звідси знаходимо рівняння осмотичного тиску:

| (8.1.2) |
Примітка: молярна концентрація речовини зовсім не обов'язково дорівнює молярній концентрації осмотично-активних часток (яку називають ще осмотичною концентрацією або осмомолярністю).
Солі, такі як NaCl або KCl у водному розчині повністю дисоціюють, і їх осмотична концентрація у два рази перевищує молярну концентрацію. Молекула CaCl2 дисоціює у водяному розчині на три части, тому її осмотична концентрація буде перевищувати молярну у три рази.
| Основне рівняння | Вид роботи | Фактор інтенсивності | Фактор ємності |
| A = Fs | Переміщення тіла | F-сила | s -переміщення |
| A = p | Стиснення газу | p -тиск газу | V -об’єм |
| A = p | Зміна концентрації | p -осмотичний тиск | q- заряд |
| A = jq | Перенос заряду | j -потенціал | q -заряд |
| A = mm | Хімічна реакція | m -хімічний потенціал | m -кількість речовини |
Види роботи:Механічна, розширення газу, осмотична, електрична.
Осмотична робота й енергія
Осмотична робота - це робота, яку треба зробити, щоб збільшити концентрацію речовини в даному розчині. Таку роботу буде робити гіпотетичний пристрій, зображений на 8.2.1. у третьому стовпчику. Він складається з циліндру, в якому роль поршня виконує пластинка з "напівпроникної речовини". Крізь пластинку може вільно проходити розчинник, але не можуть проходити молекули (іони) розчиненої сполуки.
Приклавши до такої пластинки тиск p, можна стиснути об'єм розчину з вихідного V1 до кінцевого V2; робота, яка буде зроблена, як й у випадку стиску газу, дорівнюватиме:
 (8.2.2.)
(8.2.2.)
Осмотична робота при переносі іонів або нейтральних молекул крізь мембрану
При перенесенні іонів (молекул) крізь мембрану також відбувається осмотична робота, але обчислюється вона інакше, оскільки при цьому не відбувається зміни об'єму, але відбувається зміна концентрації, а отже, зміна осмотичного тиску (див. рис. 8.2.2).
Робота при зміні тиску на величину dp при постійному об'ємі дорівнює:
При переносі іона через мембрану з одного водяного розчину, де його концентрація дорівнює С1, в інший водяний розчин, де концентрація іона дорівнює С2, відбувається робота рівна:
7. Електрична робота Нагадаємо деякі відомості з електростатики. Переміщення заряду q = ze в електричному полі з напруженістю E обумовлено дією на заряд сили F = ze. Переміщення заряду на відстань dx у напрямку, який є протилежним діючої на нього силі, зажадає здійснення роботи, яка дорівноє:
Напруженість поля пов'язана з градієнтом потенціалу в напрямку поля (вісь X) рівнянням:
З останніх двох рівнянь одержуємо рівняння роботи, затрачуваної на переміщення іона в електричному полі:
Таким чином, робота, затрачувана на перенос іона із зарядом ze з середовища з потенціалом j1 у середовище з потенціалом j2 дорівнює:
де e - елементарний заряд, тобто заряд протона в Кулонах, а z - безрозмірний заряд, іноді називаний валентністю іона. Величина z для іонів Na+, K+, Cl-, Ca2+ й Fe3+ становить відповідно 1, 1, -1, 2 й 3. |
Робота при перенесенні зарядів через мембрану
Робота, яка необхідна для переміщення m кіломолів іонів з областю з потенціалом j1 в область із потенціалом j2, дорівнює:
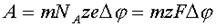
| (8.2.9) |
де NA - число Авогадро, а F - число Фарадея (заряд у Кулонах одного кіломоля одновалентних іонів).
8. Між різними ділянками живої клітини й оточуючим середовищем можуть існувати різниці потенціалів, тобто електричний потенціал всередині клітини відрізняється від зовнішнього. Клітинний мембранний потенціал визначають, як різницю потенціалів між внутріклітинним вмістом й оточуючим середовищем. Ця різниця потенціалів має назву потенціалу спокою. Якщо зовнішній потенціал прийняти за 0, то внутрішній складає від -50 до -90 мВ, в залежності від виду клітин.
Причиною виникнення потенціалу спокою є наявність градієнту концентрацій іонів К+, Na+, Cl- на клітинній мембрані.
Припустимо, що мембрана сприяє проникненню лише іонів калію і заважає проникненню інших, тоді потенціал спокою буде визначатися рівноважним калієвим потенціалом і описується рівнянням Нернста:
∆j = 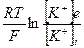
| (9.2.1.) |
де R – універсальна газова постійна, Т – абсолютна температура, F – число Фарадея, [ К+ ]0 і [ К+ ] і - зовнішня і внутрішня концентрації іонів калію відповідно.
Величина потенціалу спокою завжди декілька нижче рівноважного калієвого потенціалу, що пояснюється просочуванням всередину клітини іонів натрію, котрі приносять туди свій додатній заряд. Дифузія від’ємно заряджених іонів хлору, навпаки, сприяє збільшенню ∆j. Таким чином, внесок у формування потенціалу спокою вносять потоки всіх і
|
| Поделиться: |



