Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Подтип Mastigophora (жгутиконосцы)Стр 1 из 2Следующая ⇒
Тело жгутиконосцев разнообразной формы (яйцевидное, цилиндрическое, шаровидное, веретеновидное), покрытое плотной эластичной оболочкой - пелликулой (эвгленовые), твердой оболочкой из клетчатки (панцирные), студенистым веществом у колониальных видов или элементарной мембраной (у немногих видов, образующих псевдоподии). Минеральный скелет, за редким исключением, отсутствует. Размеры жгутиконосцев варьируют от 1 – 2 мкм до нескольких миллиметров. Органеллы передвижения - цитоплазматические нитевидные жгутики, размещенные на переднем конце тела. Число жгутиков различно (чаще один или два жгутика, реже 4,8 и более). У полимастигин жгутики покрывают большую часть поверхности клетки. Кроме постоянных жгутиков могут быть и псевдоподии (Rhizomastigida). Цитоплазма жгутиковых делится на экто- и эндоплазму. В цитоплазме находятся органоиды, обеспечивающие локомоторную функцию жгутиков, выделительную и осморегуляторную функции (сократительные вакуоли, характерны для свободноживущих пресноводных форм), разнообразные включения: жироподобные вещества, гликоген, крахмал. Зерна парамила и крахмала характерны для растительных жгутиконосцев (рисунок 3). Большинство жгутиконосцев - одноядерные организмы, существуют двухядерные (Lamblia) и многоядерные виды. У видов Opalina число ядер может быть более сотни. Способы питания жгутиковых разнообразны: голозойное (анимальное), сапрофитное, голофитное, миксотрофное. Состав пищи гетеротрофов: бактерии, водоросли, жидкая органическая пища. Пищеварительные органеллы: пищеварительная вакуоль, клеточный рот и глотка. Последние характерны для некоторых жгутиковых. Растительные жгутиконосцы обладают хромотофорами, содержащими хлорофилл, и способны к фотосинтезу. Среди них встречаются одиночные и колониальные формы. Эвгленовые и панцирные содержат светочувствительные органеллы - стигмы (глазные пятна). Гетеротрофные жгутиконосцы (зоомастигины) хроматофоров не содержат. Среди них обычны мелкие бесцветные формы с 1-3 жгутиками -обитатели загрязненных водоемов и почв разных типов, одиночные или колониальные воротничковые жгутиконосцы, многожгутиковые формы. У многих жгутиконосцев наблюдается бесполое размножение продольным делением. Половое размножение известно для колониальных растительных жгутиковых и некоторых одиночных форм.
Многие жгутиконосцы - обитатели моря, значительная часть которых входит в состав планктона. Видовой состав жгутиконосцев пресных водоемов богат и зависит от степени загрязненности вод органическими веществами. Среди зоомастигин много паразитических форм, возбудителей заболеваний растений, животных и человека (рисунок 4). Средой обитания паразитических форм могут быть различные органы и ткани: кишечник, кровяное русло, кожа. Наиболее патогенными видами для человека являются: Trypanosoma qambiense - возбудитель сонной болезни, Leishmania donovani - возбудитель висцерального лейшманиоза ("кала-азар"), Leishmania tropica - возбудитель восточной язвы и др. Переносчиками этих парази тических жгутиконосцев являются различные кровососущие насекомые, в организмах которых проходит часть жизненного цикла простейших. T. crusi вызывает болезнь Чагаса, поражающую людей в Америке. Паразиты живут в кровяном русле и в клетках внутренних органов человека и в кишечнике клопов-кровососов. Заражение человека происходит при попадании экскрементов клопов, при наличии в них активной фазы возбудителя, в ранки и глубокие ссадины на открытых участках кожи рук. Болезни домашних животных: нагана рогатого скота, су-аура, случная болезнь лошадей - вызываются различными видами трипаносом: Т. brucei, Т. evansi, Т. equiperdum. Случная болезнь развивается без насекомых- переносчиков. Передача паразитов происходит при случке лошадей.
Рисунок 3 – Жгутиковые: A- Euglena oxyuris; Б- Euglena viridis: 1-жгутик, 2- резервуар сократительной вакуоли. 3- сократительная вакуоль, 4-хроматофоры. несущие хлорофилл. 5- ядро, 6- парамиловые зерна. 7- глазок; В-колония вольвокса (V. globator) с половыми клетками: Г- колония вольвокса (Г aureus) в процессе бесполого размножения: 1- макрогамета. 2- микрогаметы. 3-дочерние колонии
Рисунок 4 – Паразитические жгутиконосцы:. А - схема строения жгутиконосца p. Trichomonas: 1- жгутики. 2- ундулирующая мембрана, 3-базальное тельце (кинетосома). 4- ядро, 5- аксостиль; Б- паразитирующий в кишечнике человека Trichomonas hominis: В- трипаносома (Г. vittatae) из крови черепахи: I- кинетопласт. 2- ядро. 3- ундулируюшая мембрана. 4- жгутик
Тип Sporozoa (споровики) Споровики - исключительно паразитические простейшие, разделенные на три группы: 1) грегарины - внеклеточные паразиты кольчатых червей и членистоногих, обитающие в кишечнике, полости тела и половых железах; 2) кокцидии - внутриклеточные паразиты позвоночных и беспозвоночных животных, обитающие в клетках кишечника, почек, печени, мозга и т.д. 3) кровяные споровики (гемоспоридии) - паразиты крови млекопитающих, птиц и рептилий, обитающие в клетках кровеносных сосудов печени, в эритроцитах. Споровики имеют разнообразную форму тела: продолговатую, червеобразную (с наличием или отсутствием трехчленистости) и сферическую у грегарин. округлую, овальную, серповидную у кокцидий и кровяных споровиков. Форма тела и размеры споровиков изменяются на разных этапах жизненного цикла (рисунок 5). Жизненный цикл протекает с чередованием поколений и с переходами паразитов из организма хозяина в окружающую среду и обратно (грегарины. кокцидии), а также со сменой хозяев (токсоплазмы, кровяные споровики). Сложность жизненного цикла и упрощение организации споровиков объясняется их переходом к паразитическому существованию. Органеллы активного передвижения у споровиков отсутствуют, за исключением жгутиков у мужских гамет. Снаружи тело споровиков покрыто пелликулой. Цисты, поступающие в окружающую среду, одеты плотной оболочкой (рисунок 5, 6). У кровяных споровиков такая оболочка отсутствует, так как в их жизненном цикле нет контактов с внешней средой (рисунок 7). У грегарин имеется система мионем. обеспечивающая сокращение и выпрямление тела. Многие кишечные грегарины способны к скользящим движениям, их размеры 10- 15 мкм. отдельные виды достигают 16 мм. У споровиков нет органелл захвата пищи, пищеварительных и сократительных вакуолей. Питательные вещества поступают через микропоры (ультроцитостом) у мерозоитов кокцидий и растущих шизонтов кровяных споровиков или осмотически, всасываясь всей поверхностью тела,- у грегарин. В цитоплазме споровиков присутствуют общеклеточные органоиды- митохондрии, аппарат Гольджи и др. Дыхание споровиков происходит за счет ферментативного расщепления гликогена. Все вегетативные стадии споровиков представлены гаплофазой. Зигота – единственная диплоидная стадия, которая в виде ооцисты (споры) может передаваться следующему хозяину (грегарины и кокцидии).
Рисунок 5 – Цикл развития грегарины Stylocepalus longicollis:
1- спорозоит. вышедший из ооцисты. 2- растущая грегарина. 3- сизигий (две объединившиеся грегарины). 4- циста с двумя грегаринами, 5- деление ядер. 6-образование гамет. 7- гаметы (мужские - продолговатые, женские - округлые). 8- копуляция гамет. 9- зигота, выделившая оболочку (ооциста). 9-15 - последовательные стадии спорогонии (три деления ядер, первое- редукционное) с образованием восьми спорозоитов
Рисунок 6 – Цикл развития кокцидий рода Eimeria:
I-III - последовательные поколения шизогонии, IV - гаметогония. V - спорогония: 1- спороюиты. 2.3 - развитие шизонта. 4 - шизонт. распавшийся на мерозоиты. 5- развитие макрогамет. 6-7 - развитие микрогамет. 8 - ооциста. 9 - ооциста. приступающая к спорогонии. 10.11 - развитие споробластов. 12 - зрелые ооци-сты (в каждой из четырех спор по два спорозоита)
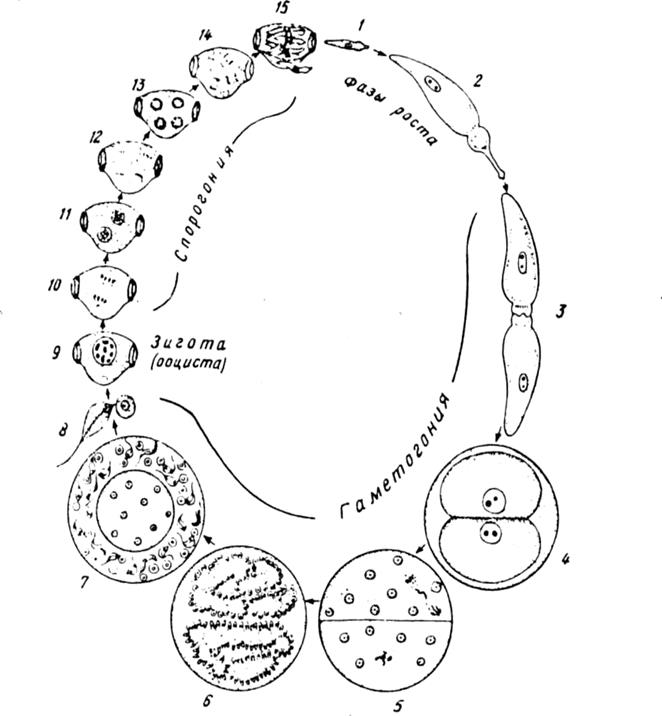
Рисунок 7 – Цикл развития малярийного плазмодия рода Plasmodium:
1 -спорозоиты, 2- 4-шизогония в печени (конечный этап - формирование мерозоитов из шизонтов) 5-10- эритроцитарная шизогония (конечный этап - формирование мерозоитов в эритроцитах). 11-12- развитие макрогамонтов, 11 а. 12а - развитие микрогамонтов. 13-макрогамета: 14,15- развитие микрогамет, 16- копуляция. 17-20 - развитие зиготы -оокинеты (конечный этап - формирование из оокинеты ооцисты на наружной стенке кишечника комара), 21-24 - дальнейшее развитие ооцисты, 24, 25 - миграция спорозоитов из ооцисты в слюнные железы комара Кровяные споровики передаются насекомыми-переносчиками промежуточному хозяину на стадии сформировавшегося спорозоита (рисунок 7). Споровики размножаются бесполым и половым способом. Бесполое размножение происходит при множественном делении ядра спорозоита или шизонта (шизогония), а также при эндодиогении (известна у кокцидий) в несколько циклов. Бесполое размножение кровяных споровиков сопровождается клеточной и эритроцитарной шизогонией с формированием нескольких поколений мерозоитов. Процесс образования половых особей называется гаметогонией, при которой число образующихся ядер во много раз больше, чем при шизогонии. После образования зиготы наблюдается спорогония, протекающая во внешней среде (грегарины и кокцидии) или в организме окончательного хозяина (токсоплазмы, кровяные споровики). Все стадии размножения способствуют быстрому увеличению численности паразитов. Многие споровики, являясь возбудителями тяжелых заболеваний домашних животных и человека, имеют большое практическое значение. К ним относятся кокцидии из рода Еimeria - возбудители кокцидоза кроликов, цыплят, крупного рогатого скота, карповых рыб. Для человека опасны представители рода Plasmodium, вызывающие различные виды лихорадок (малярия). Переносчиками заболевания являются комары p. Anopheles.
|
|||||||
|
|
Последнее изменение этой страницы: 2017-01-24; просмотров: 980; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.36.141 (0.011 с.) |