Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Судьба всосавшихся моносахаридовСтр 1 из 6Следующая ⇒
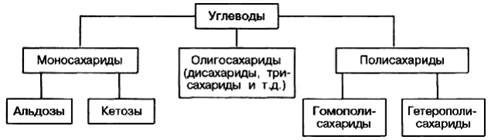
ХИМ ПРИРОДА УГЛЕВОДОВ По химической природе углеводы – полиоксикарбонильные соединения: их углеродный скелет несёт какую-либо карбонильную группу (альдегидную, кетонную, карбоксильную) и несколько гидроксильных групп. Общую формулу многих углеводов можно представить в виде Cm (H2O)n, т.е. углерод + вода (отсюда название – углеводы). Кроме того, существует множество углеводов, включающих различные группы (напр., аминогруппу NH2). У простых углеводов – моносахаридов – углеродная цепь может содержать 3 углеродных атома (триозы), 4 (тетрозы), 5 (пентозы), 6 (гексозы) и т.д. Пентозы рибоза и дезоксирибоза входят в состав нуклеотидов и нуклеиновых кислот. Гексозы глюкоза, фруктоза и галактоза играют важную роль в обеспечении всех организмов энергией. В растениях моносахариды образуются в процессе фотосинтеза и служат также исходными веществами для биосинтеза более сложных углеводов, аминокислот и других соединений. КЛАССИФИКАЦИЯ.ФУНКЦИИ. По классификации, углеводы подразделяются на три основные группы: моносахариды, олигосахариды и полисахариды. Моносахариды как производные многоатомных спиртов, содержащие карбонильную (альдегидную или кетонную) группу. Если карбонильная группа находится в конце цепи, то моносахарид представляет собой альдегид и называется альдозой; при любом другом положении этой группы моносахарид является кетоном и называется кетозой. Простейшие представители моносахаридов – триозы: глицеральдегид и
диоксиацетон. При окислении первичной спиртовой группы трехатомного спирта – глицерола – образуется глицеральдегид (альдоза), а окисление вторичной спиртовой группы приводит к образованию диоксиацетона (кетоза).5
Олигосахариды – углеводы, молекулы которых содержат от 2 до 10 остатков моносахаридов, соединенных гликозидными связями. В соответствии с этим различают дисахариды, трисахариды и т.д. Дисахариды – сложные сахара, каждая молекула которых при гидролизе распадается на две молекулы моносахаридов. Дисахариды наряду с полисахаридами являются одними из сновных источников углеводов в пище человека и животных. По строению дисахариды –это гликозиды, в которых 2 молекулы моносахаридов соединены гликозидной связью.Среди дисахаридов наиболее широко известны мальтоза, лактоза и сахароза. Мальтоза, являющаяся α-глюкопиранозил-(1–>4)-α-глюкопиранозой, образуется как ромежуточный продукт при действии амилаз на крахмал (или гликоген), содержит 2 остатка α-D-глюкозы (название сахара, полуацетальный гидроксил которого участвует в образовании гликозидной связи, оканчивается на «ил»). Мальтоза- В молекуле мальтозы у второго остатка глюкозы имеется свободный полуацетальный гидроксил. Такие дисахариды обладают восстанавливающими свойствами. Одним из наиболее распространенных дисахаридов является сахароза – обычный пищевой сахар. Молекула сахарозы состоит из одного остатка D-глюкозы и одного остатка D-фруктозы. В отличие от большинства дисахаридов сахароза не имеет свободного полуацетального гидроксила и не обладает восстанавливающими свойствами. Гидролиз сахарозы приводит к образованию смеси, которую называют инвертированным сахаром. В этой смеси преобладает сильно левовращающая фруктоза, которая инвертирует (меняет на обратный) знак вращения правовращающего раствора исходной сахарозы. Среди природных трисахаридов наиболее известна рафиноза, содержащая остатки фруктозы, глюкозы и галактозы. Рафиноза в больших количествах содержится в сахарной свекле и во многих других растениях. В целом олигосахариды, присутствующие в растительных тканях, разнообразнее по своему составу, чем олигосахариды животных тканей. — общее название класса сложных высокомолекулярных углеводов, молекулы которых состоят из десятков, сотен или тысяч мономеров — моносахаридов. Полисахариды необходимы для жизнедеятельности животных и растительных организмов. Они являются одним из основных источников энергии, образующейся в результате обмена веществ организма. Они принимают участие в иммунных процессах, обеспечивают сцепление клеток в тканях, являются основной массой органического вещества в биосфере.
Классификация полисахаридов Выделяют гомополисахариды, состоящие из одинаковых остатков моносахаров (крахмал, гликоген, целлюлоза) и гетерополисахариды (гиалуроновая кислота, хондроитинсульфаты), включающие разные моносахара. Гомополисахариды Крахмал – гомополимер α-D-глюкозы. Находится в злаках, бобовых, картофеле и некоторых других овощах. Синтезировать крахмал способны почти все растения. Двумя основными компонентами крахмала являются амилоза (15-20%) и амилопектин (80-85%). Амилоза представляет собой неразветвленную цепь с молекулярной массой от 5 до 500 кДа, в которой остатки глюкозы соединены исключительно α-1,4-гликозидными связями. Амилопектин содержит α-1,4- и α-1,6-гликозидные связи, имеет массу не менее 1 млн Да и является разветвленной молекулой, причем ветвление роисходит за счет присоединения небольших глюкозных цепочек к основной цепи посредством α-1,6- гликозидных связей. Каждая ветвь имеет длину 24-30 остатков глюкозы, веточки возникают примерно через 14-16 остатков глюкозы в цепочке. Фунции: Биологическое значение углеводов: Злоупотребление сахарозой: Сахар – это бытовое название сахарозы, которая представляет собой дисахарид, в состав которого входят два моносахарида – глюкоза и фруктоза. Сахароза содержится во многих фруктах, овощах и ягодах. Очень высокое содержание этого вещества в сахарной свекле и сахарном тростнике, которые используются для промышленного производства сахара. Для нормального функционирования человеческому организму нужнаэнергия. Свои энергетические запасы тело пополняет за счет углеводов. В результате расщепления углеводы быстро обеспечивают энергией клетки, нуждающиеся в «подзарядке». Поскольку сахар после употребления мгновенно впитывается в кровь, он способен доставить энергию клеткам значительно быстрее, чем другие углеводы.
В процессе энергетического обеспечения организма задействован гормон инсулин, который обеспечивает доставку энергии клеткам. Этот гормон вырабатывается поджелудочной железой. Но кроме сахара в наш повседневный рацион также входят белки и жиры, которые наряду с углеводами также обеспечивают организм необходимой энергией. Это приводит к возникновению избытка энергии, постоянно высокому уровню сахара в крови, а вместе с ним и высокому уровню инсулина. Ежедневное и неограниченное употребление сладостей может впоследствии привести к тому, что клетки перестанут реагировать на действие гормона инсулина. И таким образом сахар становится причиной развития сахарного диабета второго типа. ПЕРЕВАРИВАНИЕ УГЛЕВОДОВ. Переваривание бывает: 1). Внутриклеточное (в лизосомах); 2). Внеклеточное (в ЖКТ): а). полостное (дистантное); б). пристеночное (контактное).Расщепление углеводов начинается в полости рта под действием амилазы слюны.Известны три вида амилаз, которые различаются главным образом по конечным продуктам их ферментативного действия: α-амилаза, β-амилаза и γ-амилаза. α-Амилаза расщепляет в полисахаридах внутренние α-1,4-свя-зи, поэтому ее иногда называют эндоамилазой. Молекула α-амилазы содержит в своих активных центрах ионы Са 2+ необходимые для ферментативной активности. Под действием β-амилазы от крахмала отщепляется дисахарид мальтоза, т.е. β- амилаза является экзоамилазой. Она обнаружена у высших растений, где выполняет важную роль в мобилизации резервного (запасного) крахмала. γ-Амилаза отщепляет один за другим глюкозные остатки от конца полигликозидной цепочки Переваривание углеводов в ротовой полости (полостное) В ротовой полости пища измельчается при пережѐвывании и смачивается слюной. Слюна состоит на 99% из воды и обычно имеет рН 6,8. В слюне присутствует эндогликозидаза α-амилаза (α-1,4-гликозидаза), расщепляющая в крахмале внутренние α- 1,4-гликозидные связи с образованием крупных фрагментов — декстринов и небольшого количества мальтозы и изомальтозы. Переваривание углеводов в желудке (полостное) Действие амилазы слюны прекращается в кислой среде (рН <4) содержимого желудка, однако, внутри пищевого комка активность амилазы может некоторое время сохраняться.. Переваривание углеводов в тонком кишечнике (полостное и пристеночное) В двенадцатиперстной кишке кислое содержимое желудка нейтрализуется соком поджелудочной железы (рН 7,5—8,0 за счет бикарбонатов). С соком поджелудочной железы в кишечник поступает панкреатическая α-амилаза. Эта эндогликозидаза гидролизует внутренние α-1,4-гликозидные связи в крахмале и декстринах с образованием мальтозы, изомальтозы и олигосахаридов, содержащих 3—8 остатков глюкозы, связанных α-1,4- и α- 1,6-гликозидными связями. Переваривание мальтозы, изомальтозы и олигосахаридов происходит под действием специфических ферментов - экзогликозидаз, образующих ферментативные комплексы. Эти комплексы находятся на поверхности эпителиальных клеток тонкого кишечника и осуществляют пристеночное пищеварение: Сахаразо-изомальтазный комплекс состоит из 2 пептидов, имеет доменное строение. Из первого пептида образован цитоплазматический, трансмембранный (фиксирует 9 комплекс на мембране энтероцитов) и связывающий домены и изомальтазная субъединица. Из второго - сахаразная субъединица. Сахаразная субъединица гидролизует α-1,2- гликозидные связи в сахарозе, изомальтазная субъединица - α-1,6-гликозидные связи в изомальтозе, α-1,4-гликозидные связи в мальтозе и мальтотриозе. Комплекса много в тощей кишке, меньше в проксимальной и дистальной частях кишечника. Гликоамилазный комплекс, содержит две каталитические субъединицы, имеющие небольшие различия в субстратной специфичности. Гидролизует α-1,4-гликозидные связи в олигосахаридах (с восстанавливающего конца) и в мальтозе. Наибольшая активность в нижних отделах тонкого кишечника. β-Гликозидазный комплекс (лактаза) гликопротеин, гидролизует β-1,4-гликозидные связи в лактозе. Активность лактазы зависит от возраста. У плода она особенно повышена в поздние сроки беременности и сохраняется на высоком уровне до 5-7-летнего возраста. Затем активность лактазы снижается, составляя у взрослых 10% от уровня активности, характерного для детей. Переваривание углеводов заканчивается образованием моносахаридов – в основном глюкозы, меньше образуется фруктозы и галактозы, еще меньше – маннозы, ксилозы и арабинозы.
Всасывание углеводов . Моносахариды всасываются эпителиальными клетками тощей и подвздошной кишок. Транспорт моносахаридов в клетки слизистой оболочки кишечника может осуществляться путѐм диффузии (рибоза, ксилоза, арабиноза), облегчѐнной диффузии с помощью белков переносчиков (фруктоза, галактоза, глюкоза), и путем активного транспорта (галактоза, глюкоза). Активный транспорт галактозы и глюкозы из просвета кишечника в энтероцит осуществляется симпортом с Na +. Через белок-переносчик Na + двигается по градиенту своей концентрации и переносит с собой углеводы против их градиента концентраций. Градиент концентрации Na + создаѐтся Nа + /К + -АТФ- зой. При низкой концентрации глюкозы в просвете кишечника она транспортируется в энтероцит только активным транспортом, при высокой концентрации - активным транспортом и облегчѐнной диффузией. Скорость всасывания: галактоза > глюкоза > фруктоза > другие моносахариды. Моносахариды выходят из энтероцитов в направлении кровеносного капилляра с помощью облегченной диффузии через белки-переносчики. Расщепление углеводов начинается в полости рта под действием амилазы слюны.
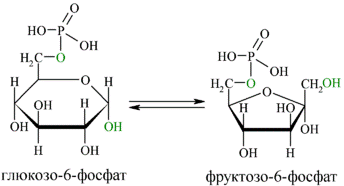
ГЛЮКОЗА 6 ФОСФАТ Образование глюкозо-6-фосфата в гексокиназной реакции сопровождается освобождением значительного количества свободной энергии системы и может считаться практически необратимым процессом. Наиболее важным свойством гексокиназы является ее ингибирование глюкозо-6- фосфатом, т.е. последний служит одновременно и продуктом реакции, и аллостерическим ингибитором. Фермент гексокиназа способен катализировать фосфорилирование не только D- глюкозы, но и других гексоз, в частности D-фруктозы, D-маннозы и т.д. В печени, кроме гексокиназы, существует фермент глюкокиназа, который катализирует фосфорилирование только D-глюкозы. В мышечной ткани этот фермент отсутствует. Второй реакцией гликолиза является превращение глюкозо-6-фосфата под действием фермента глюкозо-6-фосфат-изомеразы во фруктозо-6-фосфат:
Эта реакция протекает легко в обоих направлениях, и для нее не требуется каких-либо кофакторов.Третья реакция катализируется ферментом фосфофруктокиназой; образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второй молекулы АТФ:14 Данная реакция аналогично гексокиназной практически необратима, протекает в присутствии ионов магния и является наиболее медленно текущей реакцией гликолиза. Фактически эта реакция определяет скорость гликолиза в целом. Фосфофруктокиназа относится к числу аллостерических ферментов. Она ингибируется АТФ и стимулируется АМФ. При значительных величинах отношения АТФ/АМФ активность фосфофруктокиназы угнетается и гликолиз замедляется. Напротив, при снижении этого коэффициента интенсивность гликолиза повышается. Так, в неработающей мышце активность фосфофруктокиназы низкая, а концентрация АТФ относительно высокая. Во время работы мышцы происходит интенсивное потребление АТФ и активность фосфофруктокиназы повышается, что приводит к усилению процесса гликолиза. Четвертую реакцию гликолиза катализирует фермент альдолаза. Под влиянием этого фермента фруктозо-1,6-бисфосфат расщепляется на две фосфотриозы: Эта реакция обратима. В зависимости от температуры равновесие устанавливается на различном уровне. При повышении температуры реакция сдвигается в сторону большего образования триозофосфатов (дигидро-ксиацетонфосфата и глицеральдегид-3-фосфата). Пятая реакция – это реакция изомеризации триозофосфатов. Катализируется ферментом триозофосфатизомеразой:
ПУТИ УТИЛИЗАЦИИ ГЛЮКОЗЫ А)Пентозный цикл Пентозный цикл совокупность ферментативных реакций, в результате которых в организме происходит окислительный распад глюкозы с образованием пентоз, взаимопревращение углеводов с длиной цепочки от пяти до семи углеродных атомов, образование восстановленного никотинамидадениндинуклеотидфосфата и углекислого газа; участвует во многих процессах обмена веществ и энергии.
1 — глюкозо-6-фосфатдегидротеназа; Различают 3 этапа пентозного цикла, каждый этап включает несколько реакций. 1 этап – окислительная фаза – осуществляется дегидрогеназно-декарбоксилазной системой. Эта фаза начинается с дегидрирования 6 молекул гл-6-ф с помощью гл-6-фДГ (НАДФ). При этом образуются восстановленные НАДФН2 и фосфоглюконолактон, который снова дегидрируется НАДФ-зависимым ферментом и декарбоксилируется и через ряд промежуточных реакций превращается в пентозо-5-фосфат. 2 этап – катализируется изомеразами. На этом этапе происходит изомеризация пентозофосфатов. 3 этап – участвуют ферменты структурной перестройки углеводов. Образуются углеводы с различным количеством углеродов, в этих реакциях участвуют транскетолазы (Ко – вит В1). В качестве промежуточных продуктов ПФЦ образуются 2 молекулы ФГА и 4 молекулы фр-6-ф. Эти продукты могут включаться в гликолиз, а в основном 2 ФГА образуют фр-6-ф, и количество фр-6-ф увеличивается до 5. 5 молекул фр-6-ф изомеризуются до гл-6-ф. Гл-6-ф может снова включиться в ПФЦ, для этого надо добавить еще одну молекулу гл-6-ф. Значение пентозофосфатного пути окисления глюкозы: 1. За счет ПФЦ примерно 50% покрывается потребность организма в НАДФН2. НАДФН2, образованный в цикле может использоваться для следующих процессов – 1. В реакциях биосинтеза СЖК, ХС и КС, 2. В реакциях гидроксилирования при обезвреживании ксенобиотиков и некоторых эндогенных метаболитов, 3. В реакции восстановительного амминирования для обезвреживания аммиака и образование заменимых аминокислот. 4. Для восстановления окисленного глутатиона при участии ФАД-зависимого фермента ГР. 5. Для работы НАДФН-оксидазной системы клеток крови. 2. Пентозы, являющиеся промежуточными продуктами пентозного цикла, могут быть использованы для образования НК, Ко, НТФ, ГАГов. 3. ПФЦ – это путь перехода избытка пентоз в глюкозу, а затем в гликоген Б)Гликолиз Глико́лиз (фосфотриозный путь, или шунт Эмбдена — Мейерхофа, или путь Эмбдена-Мейергофа-Парнаса) — ферментативный процесс последовательного расщепления глюкозы в клетках, сопровождающийся синтезомАТФ. Гликолиз при аэробных условиях ведёт к образованию пировиноградной кислоты (пирувата), гликолиз в анаэробных условиях ведёт к образованию молочной кислоты (лактата). Гликолиз является основным путёмкатаболизма глюкозы в организме животных. представляет собой 10 последовательных реакций, каждая из которых катализируется отдельным ферментом. В любом гликолизе можно выделить 2 этапа:
Процесс гликолиза условно можно разделить на два этапа. Первый этап, протекающий с расходом энергии 2-х молекул АТФ, заключается в расщеплении молекулы глюкозы на 2 молекулы глицеральдегид-3-фосфата. На втором этапе происходит НАД-зависимое окисление глицеральдегид-3-фосфата, сопровождающееся синтезом АТФ. Сам по себе гликолиз является полностью анаэробным процессом, то есть не требует для протекания реакций присутствия кислорода. Результатом гликолиза является превращение одной молекулы глюкозы в две молекулы пировиноградной кислоты (ПВК) и образование двух восстановительных эквивалентов в видекофермента НАД∙H. Полное уравнение гликолиза имеет вид: Глюкоза + 2НАД+ + 2АДФ + 2Фн = 2НАД∙Н + 2ПВК + 2АТФ + 2H2O + 2Н+. При отсутствии или недостатке в клетке кислорода пировиноградная кислота подвергается восстановлению до молочной кислоты, тогда общее уравнение гликолиза будет таким: Глюкоза + 2АДФ + 2Фн = 2лактат + 2АТФ + 2H2O. Таким образом, при анаэробном расщеплении одной молекулы глюкозы суммарный чистый выход АТФ составляет две молекулы, полученные в реакциях субстратного фосфорилирования АДФ.
Первой реакцией гликолиза является фосфорилирование молекулы глюкозы, происходящее при участии тканеспецифичного фермента гексокиназы с затратой энергии 1 молекулы АТФ; образуется активная форма глюкозы — глюкозо-6-фосфат (Г-6-Ф):
Для протекания реакции необходимо наличие в среде ионов Mg2+, с которым комплексно связывается молекула АТФ. Эта реакция необратима и является первой ключевой реакцией гликолиза. Фосфорилирование глюкозы преследует две цели: во-первых, из-за того что плазматическая мембрана, проницаемая для нейтральной молекулы глюкозы, не пропускает отрицательно заряженные молекулы Г-6-Ф, фосфорилированная глюкоза оказывается запертой внутри клетки. Во-вторых, при фосфорилировании глюкоза переводится в активную форму, способную участвовать в биохимических реакциях и включаться в метаболические циклы. Печёночный изофермент гексокиназы — глюкокиназа — имеет важное значение в регуляции уровня глюкозы в крови. В следующей реакции (2) ферментом фосфоглюкоизомеразой Г-6-Ф превращается во фруктозо-6-фосфат (Ф-6-Ф):
Энергия для этой реакции не требуется, и реакция является полностью обратимой. На данном этапе в процесс гликолиза может также включаться путём фосфорилирования и фруктоза. Далее почти сразу друг за другом следуют две реакции: необратимое фосфорилирование фруктозо-6-фосфата (3) и обратимое альдольное расщепление образовавшегося фруктозо-1,6-бифосфата (Ф-1,6-бФ) на две триозы (4). Фосфорилирование Ф-6-Ф осуществляется фосфофруктокиназой с затратой энергии ещё одной молекулы АТФ; это вторая ключевая реакция гликолиза, её регуляция определяет интенсивность гликолиза в целом.
Альдольное расщепление Ф-1,6-бФ происходит под действием альдолазы фруктозо-1,6-бифосфата:
В результате четвёртой реакции образуются дигидроксиацетонфосфат и глицеральдегид-3-фосфат, причём первый почти сразу под действием фосфотриозоизомеразы переходит во второй (5), который и участвует в дальнейших превращениях:
Каждая молекула глицеральдегидфосфата окисляется НАД+ в присутствии дегидрогеназы глицеральдегидфосфата до 1,3-дифосфоглицерата (6):
Далее с 1,3-дифосфоглицерата, содержащего макроэргическую связь в 1 положении, ферментом фосфоглицераткиназой на молекулу АДФ переносится остаток фосфорной кислоты (реакция 7) — образуется молекула АТФ:
Это первая реакция субстратного фосфорилирования. С этого момента процесс расщепления глюкозы перестаёт быть убыточным в энергетическом плане, так как энергетические затраты первого этапа оказываются компенсированными: синтезируются 2 молекулы АТФ (по одной на каждый 1,3-дифосфоглицерат) вместо двух потраченных в реакциях 1 и 3. Для протекания данной реакции требуется присутствие в цитозоле АДФ, то есть при избытке в клетке АТФ (и недостатке АДФ) её скорость снижается. Поскольку АТФ, не подвергающийся метаболизму, в клетке не депонируется а просто разрушается, то эта реакция является важным регулятором гликолиза. Затем последовательно: фосфоглицеролмутаза образует 2-фосфоглицерат (8):
Енолаза образует фосфоенолпируват (9):
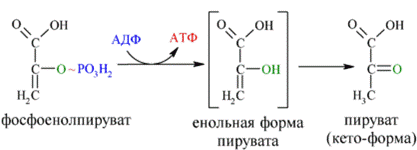
И наконец происходит вторая реакция субстратного фосфорилирования АДФ с образованием енольной формы пирувата и АТФ (10):
Реакция протекает под действием пируваткиназы. Это последняя ключевая реакция гликолиза. Изомеризация енольной формы пирувата в пируват происходит неферментативно. С момента образования Ф-1,6-бФ с выделением энергии протекают только реакции 7 и 10, в которых и происходит к субстратное фосфорилирование АДФ.
В)аэробное окисление глюкозы Аэробное окисление глюкозы В аэробных условиях глюкоза окисляется до СО2 и Н2О. Суммарное уравнение: С6Н12О6 + 6О2 → 6СО2+ 6Н2О + 2880 кДж/моль. Этот процесс включает несколько стадий: 1. Аэробный гликолиз. В нем происходит окисления 1 глюкозы до 2 ПВК, с образованием 2 АТФ (сначала 2 АТФ затрачиваются, затем 4 образуются) и 2 НАДН2; 2. Превращение 2 ПВК в 2 ацетил-КоА с выделением 2 СО2 и образованием 2 НАДН2; 3. ЦТК. В нем происходит окисление 2 ацетил-КоА с выделением 4 СО2, образованием 2 ГТФ (дают 2 АТФ), 6 НАДН2 и 2 ФАДН2; 4. Цепь окислительного фосфорилирования. В ней происходит окисления 10 (8) НАДН2, 2 (4) ФАДН2 с участием 6 О2, при этом выделяется 6 Н2О и синтезируется 34 (32) АТФ. Катаболизм глюкозы без О2 идет в анаэробном гликолизе и ПФШ (ПФП). · В ходе анаэробного гликолиза происходит окисления 1 глюкозы до 2 молекул молочной кислоты с образованием 2 АТФ (сначала 2 АТФ затрачиваются, затем 4 образуются). В анаэробных условиях гликолиз является единственным источником энергии. Суммарное уравнение: С6Н12О6 + 2Н3РО4 + 2АДФ → 2С3Н6О3 + 2АТФ + 2Н2О. · В ходе ПФП из глюкозы образуются пентозы и НАДФН2. В ходе ПФШ из глюкозы образуются только НАДФН2.
А. Метаболизм фруктозы Значительное количество фруктозы, образующееся при расщеплении сахарозы, прежде чем поступить в систему воротной вены, превращается в глюкозу уже в клетках кишечника. Другая часть фруктозы всасывается с помощью белка-переносчика, т.е. путём облегчённой диффузии. Метаболизм фруктозы (рис. 7-69) начинается с реакции фосфорилирования (реакция 1), катализируемой фруктокиназой с образованием фруктозо-1-фосфата. Фермент обнаружен в печени, а также в почках и кишечнике. Этот фермент обладает абсолютной специфичностью, поэтому, в отличие от глкжокиназы, инсулин не влияет на его активность. Последнее обстоятельство объясняет, почему уровень выведения фруктозы в моче у больных сахарным диабетом и здоровых не отличается. Фруктозе-1-фосфат не может превращаться во фруктозо-6-фосфат из-за отсутствия соответствующего фермента. Вместо этого фруктозо-1-фосфат далее расщепляется фруктозо-1-фосфатальдолазой (альдолаза В) на глицеральдегид и дигидроксиацетон-3-фосфат (реакция 2). Последний является промежуточным продуктом гликолиза и образуется в ходе реакции, катализируемой фруктозо-1,6-бисфосфосфатальдолазой (альдолаза А). Глицеральдегид может включаться в гликолиз после его фосфорилирования с участием АТФ (реакция 3). Две молекулы триозофосфатов либо распадаются по гликолитическому пути, либо конденсируются с образованием фруктозо-1,6-бисфосфата и далее участвуют в глюконеогенезе (реакции 8, 7, 5, 9). Фруктоза в печени включается главным образом во второй путь. Часть дигидроксиацетон-3-фосфата может восстанавливаться до глицерол-3-фосфата и участвовать в синтезе триацилглицеролов. Следует отметить, что включение фруктозы в метаболизм через фруктозо-1-фосфат минует стадию, катализируемую фосфофруктокиназой (реакция 6), которая является пунктом метаболитического
Рис. 7-69. Метаболизм фруктозы. а - превращение фруктозы в дигидроксиацетон-3-фосфат и глицеральдегид-3-фосфат; б - путь включения фруктозы в гликолиз и глюконеогенез; в - путь включения фруктозы в синтез гликогена. контроля скорости катаболизма глюкозы. Этим обстоятельством можно объяснить, почему увеличение количества фруктозы ускоряет в печени процессы, ведущие к синтезу жирных кислот, а также их этерификацию с образованием триацилглицеролов. Таблица 7-5. Нарушения метаболизма фруктозы
в метаболизм. Поэтому происходит торможение распада гликогена на стадии образования глюкозо-1 -фосфата, в результате чего развивается гипогликемия. Как следствие, ускоряется мобилизация липидов и окисление жирных кислот. Следствием ускорения окисления жирных кислот и синтеза кетоновых тел, замещающих энергетическую функцию глюкозы, может быть метаболический ацидоз (см. раздел 8), так как кетоновые тела являются кислотами и при высоких концентрациях снижают рН крови. Результатом торможения гликогенолиза и гликолиза является снижение синтеза АТФ. Кроме того, накопление фосфорилированной фруктозы ведёт к нарушению обмена неорганического фосфата и гипофосфатемии. Для пополнения внутриклеточного фосфата ускоряется распад адениловых нуклеотидов. Продукты распада этих нуклеотидов включаются в катаболизм, проходя стадии образования гипоксантина, ксантина и, наконец, мочевой кислоты. Повышение количества мочевой кислоты и снижение экскреции уратов в условиях метаболического ацидоза проявляются в виде гипер-урикемии. Следствием гиперурикемии может быть подагра даже в молодом возрасте (см. раздел 10). В. Метаболизм галактозы Галактоза образуется в кишечнике в результате гидролиза лактозы. Чтобы превратить галактозу в глюкозу, необходимо изменить оптическую конфигурацию Н- и ОН-групп С4 атома в галактозе, т.е. провести реакцию эпимеризации. Эта реакция в клетке возможна только с УДФ-произ-водным галактозы. УДФ-галактоза образуется из УДФ-глюкозы (метаболит в синтезе гликогена) в ходе реакции, катализируемой уридилфосфат-4-эпимеразой (рис. 7-70, 7-71). Однако включению галактозы в описанную реакцию эпимеризации предшествует её фос-форилирование с образованием галактозо-1-фосфата (реакция 1 на рис. 7-70). Далее галактозо-1-фосфат замещает остаток глюкозы в УДФ-глюкозе с образованием УДФ-галактозы (реакция 2), т.е. прямая реакция фосфорилированной галактозы с УТФ не происходит. Реакцию 2 можно рассматривать как перенос уридильного остатка с УДФ-глюкозы на галактозу, поэтому фермент назван галактозо-1 -фосфатуридилтрансферазой (ГАЛТ). Затем галактоза в составе нуклеотида включается в реакцию эпимеризации, в которой участвует эпимераза - NAD-зависимый фермент, катализирующий окисление и восстановление галактозы по С4углеродному атому (реакция 3). Эпимераза может работать и в другом направлении, преобразуя УДФ-глюкозу в УДФ-галактозу. Эта обратная эпимеризация важна для синтеза галактозильных остатков в гликолипидах и гликопротеинах. Кроме того, галактоза необходима для синтеза лактозы в грудных железах. В период лактации галактоза не является незаменимым компонентом пищи, так как может образовываться из глюкозы. Глюкозо-1-фосфат, образованный в реакции 2, может включаться в разные метаболические пути: 1) синтез гликогена после реакции с УДФ и образования УДФ-глюкозы; 2) превращение в печени
Рис. 7-70. Обмен галактозы.
Рис. 7-71. Реакция эпимеризации УДФ-глюкозы в УДФ-галактозу. в свободную глюкозу и поддержание её концентрации в крови; 3) катаболизм, сопряжённый с синтезом АТФ, и т.д. (см. рис. 7-70). Таблица 7-6. Нарушения обмена галактозы
Таблица 7-7. Некоторые варианты генетического дефекта ГАЛТ
обмене (фосфоглюкомутазы, глюкозо-6-фосфатдегидрогеназы). Известно несколько форм галактоземии, причиной которой является недостаточность ГАЛТ (табл. 7-7).
|
|||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-01-24; просмотров: 1102; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.191.195.110 (0.089 с.) |