
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Тема 11. 3. Строение и биосинтез гормонов
1. Пептидные гормоны синтезируются, как и другие белки, в процессе трансляции из аминокислот. Некоторые пептидные гормоны - это короткие пептиды; например, гормон гипоталамуса тиреотропин - либерин - трипептид. Большинство гормонов передней доли гипофиза - гликопротеины. Некоторые пептидные гормоны являются продуктами общего гена (рис. 11.3). Большинство полипептидных гормонов синтезируется в виде неактивных предшественников - препрогормонов. Образование активных гормонов происходит путем частичного протеолиза. 2. Инсулин - полипептид, состоящий из двух полипептидных цепей. Цепь А содержит 21 аминокислотный остаток, цепь В - 30 аминокислотных остатков. Обе цепи соединены между собой двумя дисульфидными мостиками. Молекула инсулина содержит также внутримолекулярный дисульфидный мостик в А-цепи. Биосинтез инсулина начинается с образования неактивных предшественников, препроинсулина и проинсулина, которые в результате последовательного протеолиза превращаются в активный гормон. Биосинтез препроинсулина начинается с образования сигнального пептида на полирибосомах, связанных с эндоплазматическим ретикулумом. Сигнальный
А - ПОМК (проопиомеланокортин) синтезируется в передней и промежуточной долях гипофиза и в некоторых других тканях (кишечнике, плаценте). Полипептидная цепь состоит из 265 аминокислотных остатков; Б - после отщепления N-концевого сигнального пептида полипептидная цепь расщепляется на два фрагмента: АКТГ (39 а.к.) и β-липотропин (42-134 а.к.); В, Г, Д - при дальнейшем протеолизе происходит образование α- и β-МСГ (Меланоцитстимулирующего гормона) и эндорфинов. КППДГ - кортикотропиноподобный гормон промежуточной доли гипофиза. Процессинг ПОМК в передней и промежуточной долях гипофиза протекает по-разному, с образованием разного набора пептидов пептид проникает в просвет эндоплазматического ретикулума и направляет в ЭР растущую полипептидную цепь. После окончания синтеза препроинсулина сигнальный пептид отщепляется (рис. 11.4). Проинсулин (86 аминокислотных остатков) поступает в аппарат Гольджи, где под действием специфических протеаз расщепляется в нескольких участках с образованием инсулина (51 аминокислотный остаток) и С-пептида, состоящего из 31 аминокислотного остатка. Инсулин и С-пептид в эквимолярных количествах включаются в секреторные гранулы. В гранулах инсулин соединяется с цинком, образуя димеры и гексамеры. Зрелые гранулы сливаются с плазматической мембраной, и инсулин и С-пептид секретируются во внеклеточную жидкость в результате экзоцитоза. После секреции в кровь олигомеры инсулина распадаются. Период полураспада инсулина в плазме крови составляет 3-10 минут, С-пептида - около 30 минут. Деградация инсулина происходит под действием фермента инсулиназы в основном в печени и в меньшей степени - в почках.
Главным стимулятором синтеза и секреции инсулина является глюкоза. Секреция инсулина усиливается также некоторыми аминокислотами (особенно аргинином и лизином), кетоновыми телами и жирными кислотами. Адреналин, соматостатин и некоторые пептиды ЖКТ тормозят секрецию инсулина.
1 - синтез полипептидной цепи проинсулина; 2 - синтез происходит на полирибосомах, прикрепленных к наружной поверхности мембраны ЭР; 3 - сигнальный пептид отщепляется о завершении синтеза полипептидной цепи и образуется проинсулин; 4 - проинсулин транспортируется из ЭР в аппарат Гольджи и расщепляется на инсулин и С-пептид; 5 - инсулин и С-пептид включаются в секреторные гранулы и выделяются путем экзоцитоза (6); ЭР - эндоплазматический ретикулум; N - концевая часть молекулы; 3. Глюкагон - одноцепочечный полипептид, состоящий из 29 аминокислотных остатков. Биосинтез глюкагона происходит в α-клетках островков Лангерганса из неактивного предшественника препроглюкагона, который в результате частичного протеолиза превращается в активный гормон. Глюкоза и инсулин подавляют секрецию глюкагона; многие соединения, включая аминокислоты, жирные кислоты, нейромедиаторы (адреналин), ее стимулируют. Период полураспада гормона составляет ~5 минут. В печени глюкагон быстро разрушается под действием специфических протеаз. 4. Соматотропин синтезируется в виде прогормона в соматотрофных клетках, которые являются наиболее многочисленными в передней доле гипофиза. Гормон роста у всех видов млекопитающих представляет собой одноцепо-
чечный пептид с молекулярной массой 22 кД, состоящий из 191 аминокислотного остатка и имеющий две внутримолекулярные дисульфидные связи. Секреция гормона роста носит пульсирующий характер с интервалами в 20-30 минут. Один из самых больших пиков отмечается вскоре после засыпания. Под влиянием различных стимулов (физические упражнения, голодание, белковая пища, аминокислота аргинин) даже у нерастущих взрослых людей уровень гормона роста в крови может возрастать до 30-100 нг/мл. Регуляция синтеза и секреции гормона роста осуществляется множеством факторов. Основной стимулирующий эффект оказывает соматолиберин, основной тормозящий - гипоталамический соматостатин. 5. Иодтиронины синтезируются в составе белка - тиреоглобулина (Тг) (рис. 11.5).
ЭР - эндоплазматический ретикулум; ДИТ - дийодтиронин; Тг - тиреоглобулин; Т3 - трийодтиронин, Т4 - тироксин. Тиреоглобулин синтезируется на рибосомах, далее поступает в комплекс Гольджи, а затем во внеклеточный коллоид, где он хранится и где происходит иодирование остатков тирозина. Образование йодтиронинов происходит в несколько этапов: транспорт иода в клетки щитовидной железы, окисление йода, йодирование остатков тирозина, образование йодтиронинов, транспорт йодтиронинов в кровь Тиреоглобулин - гликопротеин, содержит 115 остатков тирозина, синтезируется в базальной части клетки и хранится во внеклеточном коллоиде, где происходит йодирование остатков тирозина и образование йодтиронинов. Под действием тиреопероксидазы окисленный йод реагирует с остатками тирозина с образованием моноиодтиронинов (МИТ) и дийодтиронинов (ДИТ). Две молекулы ДИТ конденсируются с образованием Т4, а МИТ и ДИТ - с образованием Т3. Йодтиреоглобулин транспортируется в клетку путем эндоцитоза и гидролизуется ферментами лизосом с освобождением Т3 и Т4 (рис. 11.6).
Т3 является основной биологически активной формой йодтиронинов; его сродство к рецептору клеток-мишеней в 10 раз выше, чем у Т4. В периферических тканях в результате дейодирования части Т4 по пятому углеродному атому образуется так называемая «реверсивная» форма Т3, которая почти полностью лишена биологической активности В крови йодтиронины находятся в связанной форме в комплексе с тироксинсвязывающим белком. Только 0,03% Т4 и 0,3% Т3 находятся в свободном состоянии. Биологическая активность йодтиронинов обусловлена несвязанной фракцией. Транспортные белки служат своеобразным депо, которое может обеспечить дополнительное количество свободных гормонов. Синтез и секреция иодтиронинов регулируется гипоталамо-гипофизарной системой (рис. 11.7).
1 - тиреотропин-либерин стимулирует освобождение ТТГ; 2 - ТТГ стимулирует синтез и секрецию йодтиронинов; 3, 4 - иодтиронины тормозят синтез и секрецию ТТГ Йодтиронины регулируют процессы двух типов: • рост и дифференцировку тканей; • энергетический обмен. 6. Кортикостероиды. Общим предшественником всех кортикостероидов является холестерол. Источником холестерола для синтеза кортикостероидов служат его эфиры, поступающие в клетку в составе ЛПНП или депонированные в клетке. Освобождение холестерола из его эфиров и синтез кортикостероидов стимулируются кортикотропином. Реакции синтеза кортизола происходят в разных компартментах клеток коры надпочечников (см. рис. 11.12). При синтезе кортикостероидов образуется более 40 метаболитов, различающихся по структуре и биологической активности. Основными кортикостероидами, обладающими выраженной гормональной активностью, являются кортизол - главный представитель группы глюкокортикоидов, альдостерон - основной минералокортикоид и андрогены.
На первом этапе синтеза кортикостероидов происходит превращение холестерола в прегненолон путем отщепления 6-углеродного фрагмента от боковой цепи холестерола и окисления углеродного атома С20. Прегненолон превращается в прогестерон - С21 предшественник стероидов - кортизола и альдостерона - и стероиды С19 - предшественники андрогенов. Каким именно стероидом окажется конечный продукт, зависит от набора ферментов в клетке и последовательности реакций гидроксилирования (рис. 11.8).
1 - превращение холестерола в прегненолон; 2 - образование прогестерона; 3 -гидроксилирование прогестерона (17-21-11) и образование кортизола; 4 - гидроксилирование прогестерона (21-11) и образование альдостерона; 5 - путь синтеза андрогенов Первичное гидроксилирование прогестерона 17-гидроксилазой, а затем 21- и 11-гидроксилазой приводит к синтезу кортизола. Реакции образования альдостерона включают гидроксилирование прогестерона сначала 21-гид- роксилазой, а затем 11-гидроксилазой (см. рис. 11.8). Скорость синтеза и секреции кортизола регулируется гипоталамо-гипофизарной системой по механизму обратной отрицательной связи (рис. 11.9). Стероидные гормоны транспортируются кровью в комплексе со специфическими транспортными белками. Катаболизм гормонов коры надпочечников происходит прежде всего в печени. Здесь протекают реакции гидроксилирования, окисления и
1 - стимуляция синтеза кортикотропин-либерина; 2 - кортикотропинлиберин стимулирует синтез и секрецию АКТГ; 3 - АКТГ стимулирует синтез и секрецию кортизола; 4 - кортизол тормозит секрецию АКТГ и кортиколиберина восстановления гормонов. Продукты катаболизма кортикостероидов (кроме кортикостерона и альдостерона) выводятся с мочой в форме 17-кетостероидов. Эти продукты метаболизма выделяются преимущественно в виде конъюгатов с глюкуроновой и серной кислотами. У мужчин 2/3 кетостероидов образуется за счет кортикостероидов и 1/3 - за счет тестостерона (всего 12-17 мг в сутки). У женщин 17-кетостероиды образуются преимущественно за счет кортикостероидов (7-12 мг в сутки).
|
||||||||||||
|
|
Последнее изменение этой страницы: 2017-01-24; просмотров: 548; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.143.218.146 (0.015 с.) |
 Рис. 11.3. Образование пептидных гормонов, являющихся продуктами общего гена:
Рис. 11.3. Образование пептидных гормонов, являющихся продуктами общего гена: Рис. 11.4. Схема биосинтеза инсулина в клетках поджелудочной железы:
Рис. 11.4. Схема биосинтеза инсулина в клетках поджелудочной железы: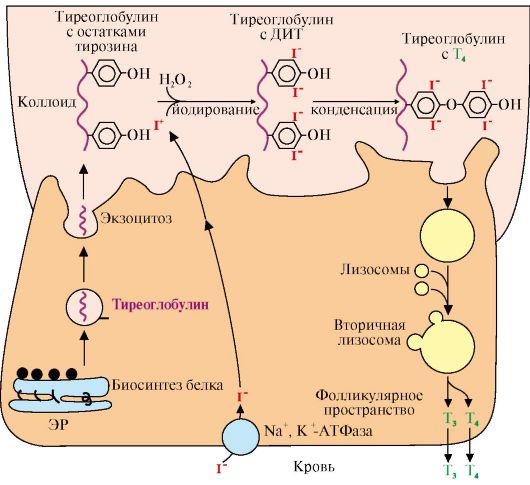 Рис. 11.5. Синтез йодтиронинов:
Рис. 11.5. Синтез йодтиронинов: Рис. 11.6. Структура гормонов щитовидной железы
Рис. 11.6. Структура гормонов щитовидной железы Рис. 11.7. Регуляция синтеза и секреции йодтиронинов:
Рис. 11.7. Регуляция синтеза и секреции йодтиронинов: Рис. 11.8. Синтез основных кортикостероидов:
Рис. 11.8. Синтез основных кортикостероидов: Рис. 11.9. Регуляция синтеза и секреции кортизола:
Рис. 11.9. Регуляция синтеза и секреции кортизола:


