Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Гормоны, как и другие сигнальные молекулы, обладают некоторыми общими свойствами. ⇐ ПредыдущаяСтр 7 из 7
Гормоно́иды(катехоламины) (гр. eidos вид) иначе парагормоны - биологически активные вещества, действующие на многие физиологические процессы в организме подобно гормонам, но образующиеся в отличие от них не в железах внутренней секреции, а в других органах и тканях; к гормоноидам относят секретин, гистамин. Классификация гормонов:
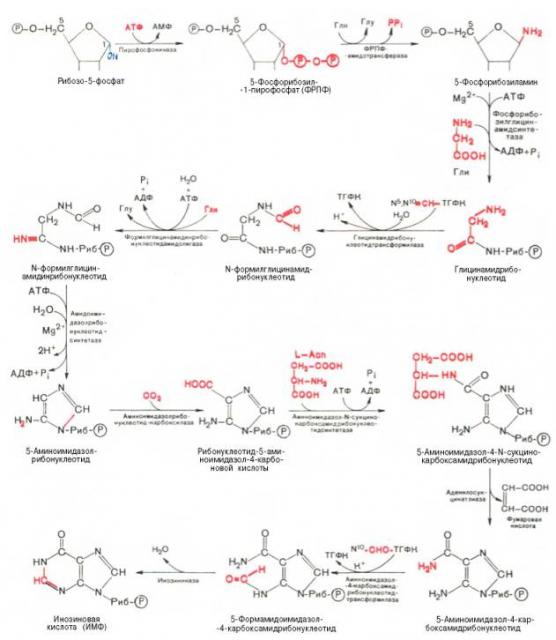
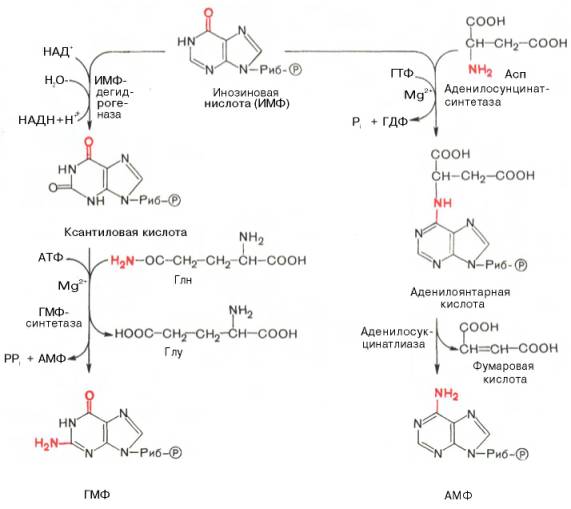
Вопрос3- биосинтез пуриновых нуклеотидов. Источники образования пуриновых оснований. Регуляция биосинтеза. Синтез пуриновых нуклеотидов осуществляется из инозинмонофосфата (ИМФ). Азотистое основание ИМФ гипоксантин превращается в аденин (первая стадия) и в гуанин (вторая стадия). Так образуются нуклеозидмонофосфаты АМФ и ГМФ, которые превращаются в нуклеозиддифосфаты АДФ и ГДФ. Процесс контролируется нуклеозидфосфаткиназами. Под действием нуклеозиддифосфаткиназ АДФ и ГДФ превращаются в АТФ и ГТФ. Нуклеозидтрифосфаты используются либо для построения РНК, либо в качестве коферментов. Преобразование рибонуклеотидов в дезоксирибонуклеотиды происходит на стации дифосфатов и катализируется нуклеозиддифосфатредуктазой.
На формильную группу формилглицинамидрибонуклеотида переносится амидная группа глутамина, и образуется формилглицинамидинрибонуклеотид. Затем пятичленное имидазольное кольцо замыкается с образованием 5-аминоимидазолрибонуклеотида, способного акцептировать СО2 с образованием 5-аминоимидазол-4-карбоновой кислоты. В последующем двухступенчатом процессе, в котором участвуют аспа-рагиновая кислота и АТФ, образуется 5-аминоимидазол-4-карбоксамид-рибонуклеотид с освобождением фумаровой кислоты. Азот аспарагиновой кислоты становится в 1-е положение будущего пуринового ядра. Последний углеродный атом пиримидинового кольца пурина вводится в виде формильного остатка, присоединяющегося к 5-NH2-группе. Наконец, отщепляется молекула воды, и образуется ИМФ. АМФ и ГМФ образуются из ИМФ, причем в синтезе обоих мононуклеотидов участвуют по два фермента. Образование ГМФ катализируют ИМФ-дегидрогеназа и ГМФ-синтетаза, образование АМФ – аденилосукцинатсинтетаза и аденилосукцинатлиаза. В ферментативном синтезе АМФ из ИМФ в качестве донора NH2-группы участвует аспарагиновая кислота. Источником энергии служит ГТФ. Промежуточный продукт – янтарная кислота. Превращение АМФ и ГМФ в соответствующие нуклеозидди- и нуклео-зидтрифосфаты также протекает в 2 стадии при участии специфических нуклеозидмонофосфат- и нуклеозиддифосфаткиназ: ГМФ + АТФ <=> ГДФ + АДФ Синтез пуриновых нуклеотидов тормозится конечными продуктами по принципу обратной связи, т.е. ингибированием переноса аминогруппы глутамина на 5-фосфорибозил-1-пирофосфат. Кроме того, избыток ГМФ в клетках оказывает аллостерическое торможение только на свой собственный синтез, не влияя на синтез АМФ; накопление АМФ подавляет свой синтез, не ингибируя синтез ГМФ.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-01-24; просмотров: 183; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.222.35.77 (0.005 с.) |