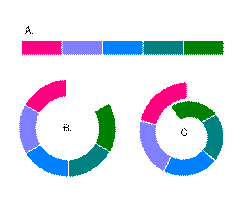Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Развитие устойчивости к антибиотикам и пестицидамСодержание книги
Поиск на нашем сайте
Развитие и распространение устойчивых к антибиотикам бактерий и устойчивых к пестицидам растений и насекомых является доказательством эволюции видов. Появление устойчивых к ванкомицину форм золотистого стафилококка и та опасность, которую они представляют для пациентов больниц, это прямой результат эволюции путем естественного отбора. Еще один пример — развитие штаммов шигеллы, устойчивых к антибиотикам из группы сульфаниламидов.Появление малярийных комаров, устойчивых к ДДТ, и развитие у австралийских популяций кроликов устойчивости к миксоматозу демонстрируют действие эволюции в условиях давления отбора при быстрой смене поколений. Естественный отбор также приводит к развитию устойчивости животных к естественным ядам, вирусам и прочим паразитам. Свежие примеры: 1) Насекомые-вредители защищаются от биологического оружия 2) Пути эволюции предопределены на молекулярном уровне (молекулярный механизм формирования у бактерий устойчивости к антибиотикам) 3) Неядовитые змеи вырабатывают устойчивость к смертоносному яду тритонов 4) Найден ген, защищающий от туберкулеза 5) Амёбы-мутанты не позволяют себя обманывать
В последнее время все большее число антиэволюционистов под давлением неопровержимых фактов, которые даже они не могут отрицать, сдают свои передовые рубежи и отступают на «запасную линию обороны», заявляя примерно следующее: «Ладно, Бог с вами, мы согласны, что микроэволюция (изменения внутри вида) возможна. Но макроэволюцию — превращение одного вида в другой — мы все равно отрицаем». В ответ на приводимые биологами доказательства превращения одного вида в другой антиэволюционисты обычно заявляют: «ну, значит, это не два разных вида, а один и тот же вид». Поскольку четкого и строго определения вида нет (и быть не может, т.к. виды постепенно эволюционируют и перетекают один в другой подчас весьма плавно и постепенно), такая аргументация оказывается принципиально непроверяемой, нефальсифицируемой, т.е. не научной. Однако у биологов есть и неопровержимые доказательства превращения друг в друга таких форм, про которые не может быть двух разных мнений: это однозначно разные виды и даже порой разные роды (хороший пример — рыбы-цихлиды африканских озер, см.: Эволюция цветного зрения у африканских цихлид шла двумя путями ). В ответ на это самые продвинутые и изощренные креационисты выдвинули концепцию «бараминов» — «сотворенных родов». Барамин — это уже самая что ни на есть распоследняя зацепка антиэволюционистов за реальность, дальше отступать им некуда — разве что в полное мракобесие и потерю рациональности (по принципу «верую, ибо абсурдно»). Согласно этой концепции, Бог сотворил большое количество разных живых существ, дав каждому возможность самостоятельно эволюционировать в неких довольно узких пределах. «Барамин» с течением времени может превратиться в пучок близкородственных видов, но дальше этого дело не пойдет никогда. «Муха останется мухой, цихлида — цихлидой, собака — собакой». Главная слабость идеи бараминов состоит в том, что из нее вытекает проверяемое следствие, которое не подверждается фактами. Следствие это состоит в том, что между «бараминами» должны быть четкие, хорошо видимые границы. Эволюционирующие виды должны «биться» о границы бараминов, никогда не выходя за них. Такие границы в природе должны быть повсеместным, всеобщим явлением, но мы этого не наблюдаем. Отдельные разрывы (хиатусы), разумеется, есть, но концепция бараминов предсказывает повсеместность таких разрывов. Однако четких, вездесущих границ между воображаемыми «бараминами» нет ни в систематике, ни в морфологии организмов, ни в сравнительной геномике, ни в биохимии, их не видно на дендрограммах сходства, построенных любыми известными методами (см. ниже в разделе «Эволюционное дерево»). И, разумеется, нет их в палеонтологической летописи, изобилующей переходными формами между крупными подразделениями (таксонами) органического мира. На практике антиэволюционисты обычно настаивают на том, что виды А и Б относятся к разным бараминам до тех пор, пока их не ткнут носом в неопровержимые доказательства родства этих видов. Тогда они говорят: «ну ладно, так и быть, значит, это один и тот же барамин». Рассмотрим пример с породами собак. «Собака осталась собакой» — любят повторять антиэволюционисты. Но генетики уже однозначно доказали, что домашние собаки происходят от волка. Значит, волк и собака — одно и то же, один и тот же барамин, говорят антиэволюционисты. А как тогда быть с шакалами и койотами, которые свободно скрещиваются с собаками и дают плодовитое гибридное потомство? Ну, значит, все они — один и тот же барамин. Мы не знаем, говорят они, какие еще виды входят в этот барамин, но если биологи докажут родство этих видов с собаками, это и будет означать, что это один и тот же барамин, а если не докажут — значит, разные барамины. Ненаучность такой аргументации очевидна. Например, сравнительная генетика доказывает родство собак с волками с абсолютно той же самой (фактически стопроцентной) степенью достоверности, что и родство их с лисами, кошками, гиенами и так далее. Антиэволюционисты ошибаются, полагая, что генетические доказательства родства быстро слабеют по мере уменьшения степени этого родства. Анализ ДНК позволяет установить отцовство — с этим даже антиэволюционисты не решаются уже спорить, но точно так же анализ ДНК позволяет установить происхождение двух особей от более далекого общего предка. Например, сейчас осуществляется активный — и весьма успешный — поиск потомков Чингисхана по всей Азии (у Чингисхана или кого-то из его ближайших предков по мужской линии произошла оригинальная мутация в У-хромосоме, что очень облегчает такой поиск) (ссылка и ссылка). Таким же способом можно установить наличие и более далекого, фактически — сколь угодно далекого общего предка, вплоть до Последнего универсального общего предка (см. выше). Анализ ДНК свидетельствует о происхождении всех ныне живущих организмов от одного общего предка. Поэтому, признав микроэволюцию, вы получаете макроэволюцию бесплатно, в виде принудительного бонуса. Не хотите — тогда доказывайте, что анализ ДНК не годится для установления отцовства. Биолога Алексея Оскольского донимал один антиэволюционист, напиравший на «отсутствие переходных форм». Оскольский спросил его: «Между тобой и твоими родителями тоже нет переходных форм, как же доказать, что именно эти люди являются твоими родителями? — «ДНК посмотреть», — не задумываясь, ответил антиэволюционист. «Ну так вот у человека и шимпанзе тоже можно ДНК посмотреть», — сказал Оскольский. Оппонента проняло.
Примеры видообразования Судя по палеонтологической летописи и по измерениям скорости мутаций, полная репродуктивная изоляция (несовместимость геномов) достигается в природе в среднем за 3 миллиона лет. Иногда больше: например, у африканских цихлид на это уходит обычно от 4 до 14 млн лет. Однако на практике биологи в большинстве случаев считают разными видами такие группы особей, которые не скрещиваются или почти не скрещиваются в природе, независимо от того, могут ли они скрещиваться в принципе. Например, тетерев и глухарь — бесспорно разные виды, хотя в принципе могут скрещиваться и давать плодовитое потомство. Другие примеры: собака, волк и шакал; очень многие виды оленей; большинство видов и даже родов африканских цихлид. Поэтому ключевым событием в видообразовании является появление не «полной» изоляции (генетической несовместимости), а фактической изоляции, когда особи из двух популяций перестают скрещиваться в природных условиях, даже если проживают на одной территории. А значит, наблюдение образования нового вида в естественных условиях в принципе возможно, но это редкое событие. Например, рассчитано, что одних только жуков появляется в среднем по два новых вида в столетие (см.: Невероятное разнообразие жуков получило эволюционное объяснение ) В то же время, в лабораторных условиях скорость эволюционных изменений может быть увеличена, поэтому ученым неоднократно удалось зафиксировать основные этапы видообразования у лабораторных животных (см. ниже: Опыты по искусственному видообразованию). Известны многие случаи видообразования посредством гибридизации и полиплоидизации у таких растений, как конопля, крапива, первоцвет, редька, капуста, а также у различных видов папоротников. У растений видообразование в природе часто происходит в результате межвидовой гибридизации и последующей полиплоидизации (удвоения хромосомного набора). Этот механизм видообразования интересен тем, что он ведет к мгновенному и автоматическому формированию полной генетической несовместимости нового вида с видами-предками. В ряде случаев видообразование у растений происходило без гибридизации и полиплоидизации (кукуруза, Stephanomeria malheurensis). Дрозофилы, также известные как плодовые мушки, входят в число наиболее изученных организмов. С 70-х годов детально изучены многие случаи видообразования у дрозофил, а отдельные этапы видообразования воспроизведены в экспериментах. Видообразование происходило в частности за счет пространственного разделения, разделения по экологическим нишам в одном ареале, изменения поведения при спаривании, дизруптивного отбора (= отбора на расхождение), а также за счет эффекта бутылочного горлышка (в ходе экспериментов founder-flush). Видообразование наблюдалось в лабораторных популяциях комнатных мух, мух Eurosta solidaginis, яблонных мух-пестрокрылок, мучных жуков, комаров и других насекомых. Зелёные водоросли и бактерии были классифицированы как отдельные виды из-за превращения одноклеточных в многоклеточные и из-за изменения строения, такого как превращение коротких отростков в длинные — все это в результате давления отбора. Видообразование также наблюдалось и у млекопитающих. Шесть случаев видообразования у домовых мышей на острове Мадейра за последние 500 лет были следствием географической изоляции, генетического дрейфа и слияния хромосом. Слияние двух хромосом — это наиболее заметное различие геномов человека и шимпанзе, а у некоторых мадейрских мышей за 500 лет было девять подобных слияний (Britton-Davidian et al., 2000).
Яблонные мухи Яблонные мухи-пестрокрылки Rhagoletis pomonella, а также паразитирующие на них наездники Diachasma являются примером наблюдаемого симпатрического видообразования (= видообразования без разделения популяции физическими барьерами). Первоначально вид обитал в восточной части США. До появления европейцев личинки этих мух развивались только в плодах боярышника. Однако с завозом в Америку яблонь (первое упоминание яблонь в Америке — 1647 год), открылась новая экологическая ниша. В 1864 году личинки Rhagoletis pomonella были обнаружены в яблоках, тем самым зафиксирована яблонная раса этого вида. За полтора века наблюдений расы очень сильно разошлись. Они почти не скрещиваются друг с другом (уровень гибридизации не превышает 4-6 %). Яблоневая раса спаривается почти исключительно на яблонях, а боярышниковая — на боярышнике, что, учитывая разное время созревания плодов, приводит к репродуктивной изоляции. Подробный «разбор полетов» яблонной мухи и ее паразитов-наездников см. в заметке: «Цепная реакция видообразования«. У пестрокрылок известно еще несколько видов-двойников, которые живут на разных видах растений — предположительно, видообразование у них протекало именно по описанной схеме (см. также: Для видообразования достаточно одного гена).
Другие хорошо документированные случаи симпатрического видообразования в природе 1) Рыбы Amphilophus citrinellus и A. zaliosus в озере Апойо в Никарагуа (Для видообразования не нужны барьеры). 2) Пальмы Howea forsteriana и H. belmoreana на острове Лорд-Хау в 580 км от восточного побережья Австралии (Эволюция без преград: ботаники нашли новое доказательство видообразования без географических барьеров). 3) Бабочки Heliconius в Центральной и Южной Америке (Зафиксирован начальный этап видообразования у тропических бабочек) 4) Трехиглая колюшка Gasterosteus aculeatus, подразделившаяся на два вида — донный и пелагический — в озерах Канады (Экспериментально подтверждено влияние видообразования на свойства экосистем). 5) Вьюрки Nesospiza acunhae и N. wilkinsi на островах Тристан-да-Кунья (Видообразование на разных островах идет параллельными путями). 6) Палочник Timema cristinae (Sandoval, 2008). 7) Стеблевой мотылек Ostrinia nubilalis. 8) Рыбы-цихлиды рода Pundamilia (Carleton et al., 2005). 9) Морская улитка Littorina saxatilis (Cruz et al., 2004). 10) Растения Mimulus lewisii и M. cardinalis. 11) Бабочка-листовертка Zeiraphera diniana (Emelianov et al., 2001). 12) Гороховая тля Acyrthosiphon pisum (Frantz et al., 2009). 13) Рыба Gambusia hubbsi (Langerhans et al., 2007). 14) Растение Anthoxanthum odoratum (Snaydon & Davies, 1976). 15) Рыбы Hypoplectrus (Puebla et al., 2007). 16) Рыбы-усачи озера Тана в Эфиопии (Sibbing et al., 1998). Опыты по искусственному видообразованию (формирование репродуктивной изоляции) Опыты по выращиванию тлей Dysaphis на «стрессовых» субстратах (малопригодных и непригодных кормовых растениях). В известных опытах, проведенных Г.Х.Шапошниковым (1961, 1965, 1966), популяция тлей Dysaphis anthrisci maicopica была пересажена с того кормового растения, на котором в природе развивается данный вид (Anthriscus nemorosa), на другое, малопригодное кормовое растение Chaerophyllum bulbosum, а затем на ранее совсем непригодное Ch. maculatum. После каждого пересаживания вначале наблюдался «кризисный» период, во время которого резко повышалась смертность и росла изменчивость. Затем популяция приспосабливалась к новым условиям: смертность и изменчивость снижались, формировалась новая «адаптивная норма», отличная от прежней. У особей преобразованной популяции обнаружилась репродуктивная несовместимость с исходной (контрольной) популяцией, не подвергавшейся воздействию стрессовых условий. Что еще более удивительно, появилась частичная совместимость с другим видом тлей, D. chaerophyllina, который является исконным обитателем растения Ch. maculatum. Результаты экспериментов Шапошникова позже были частично воспроизведены В.Б.Сапуновым (1983) в новых опытах с тлями. Опыты по дизруптивному отбору (отбору на расхождение). Почти одновременно с работами Шапошникова были опубликованы результаты экспериментов на Drosophila melanogaster по искусственному дизруптивному отбору по количественным признакам (количество щетинок). Особи с максимальными и минимальными значениями признака сохранялись, промежуточные отбраковывались. В результате между расходящимися частями популяции («много-» и «малощетинковыми» мухами) возникла частичная прекопуляционная изоляция (Thoday, Gibson, 1962). Похожие опыты были проведены на мухах Cyrtodiopsis dalmanni. У самцов этого вида глаза расположены на длинных стебельках. Данный признак поддерживается половым отбором. В эксперименте производился искусственный отбор: в одних линиях отбирались самцы с максимальным расстоянием между глазами, в других — с минимальным. Удалось вывести линии, сильно различающиеся по этому признаку. Затем были исследованы половые предпочтения самок в разных линиях. Оказалось, что в контрольных (не подвергавшихся отбору) и в «длинностебельковых» линиях самки предпочитают самцов с наибольшим расстоянием между глазами, тогда как в «короткостебельковых» линиях предпочтения самок изменились на противоположные: они стали предпочитать самцов с самыми короткими глазными стебельками. В результате между линиями возникла прекопуляционная (поведенческая) изоляция (Wilkinson, Reillo, 1994). Опыты по выращиванию Drosophila pseudoobscura на «стрессовых» средах. Опыты, сходные с экспериментами Шапошникова, позднее были проведены рядом зарубежных исследователей. Результаты получились в целом похожие, однако вскрылись некоторые новые существенные моменты, свидетельствующие о неслучайном характере изменения половых предпочтений. Рассмотрим результаты, полученные Д. Додд (Dodd, 1989) в опытах с мухами вида Drosophila pseudoobscura. Из одной природной популяции этого вида, обитавшей в каньоне Брайс (Bryce Canyon, штат Юта, США) было произведено восемь лабораторных популяций, содержавшихся изолированно. Четыре популяции выращивались на среде, основанной на крахмале, четыре другие — на мальтозе. Обе эти среды являются для данного вида малопригодными (стрессовыми). Поначалу наблюдалась высокая смертность (как и в опытах Шапошникова), и прошло несколько месяцев, прежде чем все экспериментальные популяции стабилизировались и стали «вполне здоровыми». В «крахмальных» популяциях повысилась частота встречаемости «быстрого» аллеля локуса Amy (альфа-амилазы), изменился также характер амилазной активности в средней кишке. В «мальтозных» популяциях соответствующих изменений не выявлено (Powell, Andelkovič, 1983). Однако было показано, что специфическая адаптация к стрессовым средам произошла как в «крахмальных», так и в «мальтозных» популяциях (Dodd, 1984). Затем производились эксперименты с целью выявить предпочтения при скрещивании. «Крахмальных» и «мальтозных» мух в течение одного поколения выращивали на одинаковой, стандартной (благоприятной для дрозофил) среде, а потом помещали представителей разных линий в общую камеру и наблюдали, кто с кем предпочитает спариваться. Получены статистически достоверные подтверждения того, что мухи, предки которых в течении нескольких десятков поколений росли на разных средах (предки, но не сами эти мухи!), избегают скрещиваться друг с другом. Мухи с «крахмальной» родословной достоверно предпочитали «крахмальных», «мальтозные» — «мальтозных». Опыты по выращиванию D. melanogaster в контрастных условиях. Такие же результаты получены при выращивании нескольких популяций D. melanogaster в контрастных условиях («холодно-сухо-темно» и «тепло-влажно-светло»). Здесь тоже изоляция возникла между популяциями, приспособившимися к разным условиям, тогда как между популяциями, независимо друг от друга приспособившимися к одинаковым условиям, изоляция не возникла (Kilias et al., 1980). Изоляция у дрозофил, обитающих на разных склонах «Каньона Эволюции». Явления, обнаруженные в описанных экспериментах, очевидно, имеет место и в природе. Об этом свидетельствуют результаты изучения дрозофилид, обитающих на разных склонах так называемого «Каньона Эволюции» (Evolution Canyon) на горе Кармел (Израиль). Популяции D. melanogaster, живущие на разных склонах каньона на расстоянии всего 100-400 м. друг от друга (т.е. в нескольких минутах полета) проявляют статистически достоверные предпочтения при скрещивании, избегая спариваний с обитателями другого склона. При этом никаких предпочтений между представителями разных «кланов» (изосамковых линий) с одного и того же склона не наблюдается. Условия на обоих склонах являются для дрозофил стрессовыми и сильно различаются, поскольку склон, обращенный к югу, получает в 6 раз больше солнечного излучения, чем противоположный, обращенный к северу (Korol et al., 2000). Изоляция у гаммарусов, обитающих на разных субстратах. Аналогичная ситуация наблюдается у представителей разноногих ракообразных — гаммарусов (бокоплавов), у которых подчас разные популяции одного и того же вида обитают на разных субстратах (разных видах водорослей). Эти субстраты в природе могут находиться весьма близко друг от друга, так что рачки в принципе могут свободно переплывать с одних водорослей на другие (однако, по-видимому, предпочитают этого не делать). Представители разных популяций в экспериментах демонстрируют явное предпочтение «своих» при выборе брачного партнера. Проверяют это так: в два тупика Y — образного лабиринта сажают самок, снятых с разных природных субстратов, а в непарный конец лабиринта пускают самца и смотрят, куда он поплывет. Показано, что самцы статистически достоверно предпочитают самок, живших в природе на том же субстрате, что и они сами. Быстрое развитие изоляции у лососевых рыб. Быстрое возникновение репродуктивной изоляции при расхождении по «экологическим» параметрам характерно отнюдь не только для членистоногих, о которых шла речь выше. Так, показано возникновение частичной репродуктивной изоляции между двумя популяциями нерки (Oncorhynchus nerka), которые начали расходиться от одной предковой линии не ранее, чем в 1937 г. В указанном году нерка была интродуцирована в оз. Вашингтон (штат Вашингтон, США) и вскоре образовала крупную местную популяцию во впадающей в озеро реке (Cedar River). В 1957 г. было обнаружено, что часть особей стала нереститься не в реке, а в озере. В 1992 г. было установлено, что «речные» и «озерные» формы уже представляют собой две частично изолированные популяции, немного отличающиеся морфологически. Некоторые представители «речной» популяции иногда нерестятся в озере, в тех же местах, что и «озерные» формы, однако даже в этом случае скрещивания между представителями разных популяций происходят, по-видимому, значительно реже, чем это диктуется простой вероятностью (Hendry et al., 2000). Репродуктивная изоляция в данном случае развилась самое большее за 56 лет, что соответствует примерно 13 поколениям. Все эти факты говорят о том, что репродуктивная изоляция, считающаяся одним из главных критериев вида (или даже самым главным), может быстро развиваться под действием отбора как в эксперименте, так и в природных условиях. Возникновение репродуктивной изоляции — ключевой этап видообразования. После того, как между генофондами двух популяций прекратился обмен генами, дальнейшее расхождение (дивергенция) этих популяций становится неизбежной просто в силу статистических факторов. В двух популяциях будут фиксироваться разные мутации, как нейтральные, так и полезные. Рано или поздно это неизбежно приведет к полной генетической несовместимости двух разошедшихся видов. Каждая новая мутация должна быть совместима с остальными генами в генофонде популяции, иначе отбор ее отсеет. Однако мутации, возникающие в одной из популяций, не проверяются отбором на совместимость с генофондом другой популяции. Следовательно, рано или поздно в популяции А закрепится мутация, несовместимая с генофондом популяции Б. В природе на формирование полной генетической несовместимости между двумя изолированными популяциями уходит, как правило, от 2-3 до 10-15 млн лет. Но уже задолго до этого момента разошедшие популяции считаются (и являются) двумя разными видами, потому что реально не скрещиваются в природе, хотя и сохраняют такую возможность.
Промежуточные этапы видообразования
Диаграмма, демонстрирующая варианты скрещивания соседних популяций одного вида. В третьем случае распространение форм может образовать кольцо, в результате две соседние популяции друг с другом не скрещиваются, т.е. формально являются разными видами, хотя и связаны между собой непрерывным рядом взаимно совместимых популяций.
В определении понятия биологического вида существенное значение имеет практически полная репродуктивная изоляция в природных условиях. Если видообразование происходит постепенно, в результате небольших последовательных изменений, то и сегодня мы должны наблюдать все возможные стадии видообразования, а значит и все возможные стадии репродуктивной изоляции: свободное скрещивание между популяциями, затрудненное скрещивание, снижение плодовитости потомства, бесплодное потомство, и наконец — полную генетическую изоляцию. Если ничего подобного среди современных видов обнаружить не удается, то тем самым опровергаются основы эволюционного учения, то есть это пример фальсифицируемости эволюционного учения (см.: Speciation stages). Существует множество примеров того, как разные виды могут скрещиваться в исключительных условиях. Например, у черной вороны и серой вороны разные ареалы, но на пересечении этих ареалов виды могут образовывать гибриды. Еще один пример — различные виды рыбы рода чукучанов из реки Платт, которые живут рядом, но редко скрещиваются. Неполное видообразование можно наблюдать у различных кольцевых видов: представители вида обитают вокруг гор, водоемов и т. п., при этом соседние популяции скрещиваются между собой и незначительно различаются, а крайние формы различаются значительно и не скрещиваются (см. диаграмму). Пример кольцевого вида — безлёгочная саламандра Ensatina eschscholtzii, обитающая на склонах гор близ тихоокеанского побережья Северной Америки. В зависимости от района обитания вокруг гор саламандры образуют различные формы, постепенно изменяющие свои морфологические и экологические характеристики. Крайние формы Ensatina eschscholtzii eschscholtzii и Ensatina eschscholtzii klauberi, первая из которых имеет красноватую окраску, а вторая чёрно-белую, сосуществуют в узкой зоне гор на юге Калифорнии, однако между собой не скрещиваются. Еще один пример — серебристая чайка (Larus argentatus) и клуша (Larus fuscus). Они встречаются в Англии, где их признали разными видами. Но при этом между ними существует цепочка гибридных популяций, идущая вокруг северного полюса. Другие примеры кольцевых видов — зелёная пеночка (Phylloscopus trochiloides), оленья мышь (Реromyscus maniculatus), наземная улитка (Partula olympia), дрозофила Drosophila paulistorum. У многих гибридов снижена фертильность. У гибридов землероек из рода бурозубок размножение затруднено вследствие разницы хромосомных наборов. Тот же эффект наблюдался в лаборатории при скрещивании Drosophila pseudoobscura из Юты и Калифорнии. Фертильность также снижена у гибридов лягушек из семейства бомбиновых жерлянки краснобрюхой (Bombina bombina) и жерлянки желтобрюхой (Bombina variegata). Многие другие виды могут давать бесплодное гибридное потомство. В их числе — различные земноводные (например — некоторые виды лягушек из рода Rana) и млекопитающие (например, мул — гибрид от скрещивания лошади и осла — как правило бесплоден). Гибриды гребенчатого тритона и мраморного тритона бесплодны вследствие наличия непарных хромосом. При скрещивании ряда других видов достигается оплодотворение, но позднее зародыш погибает. В качестве примера можно привести леопардовую лягушку (Rana pipiens) и лесную лягушку (Rana sylvatica) из рода настоящих лягушек. Аналогичная ситуация наблюдается у дрозофил и у некоторых растений, таких как хлопчатник обыкновенный (Gossypium hirsutum) и хлопчатник барбадосский (Gossypium barbadense). Белый медведь Конкретным примером крупномасштабных эволюционных изменений является белый медведь (Ursus maritimus), который, несмотря на родство с бурым медведем (Ursus arctos), очевидное в силу того факта, что эти виды могут скрещиваться и давать плодовитое потомство, приобрел значительные физиологические различия с бурым медведем. Эти различия позволяют белому медведю комфортно жить в условиях, в которых бурый медведь бы не выжил. В частности, белый медведь способен проплыть десятки километров в ледяной воде, сливается со снегом и не замерзает в Арктике. Всё это возможно благодаря конкретным изменениям: белая окраска способствует маскировке хищника при охоте на тюленей; полые волоски увеличивают плавучесть и сохраняют тепло; слой подкожного жира, толщина которого к зиме доходит до 10 сантиметров, обеспечивает дополнительную теплоизоляцию; удлиненная по сравнению с другими медведями шея позволяет легче держать голову над водой во время плавания; увеличенные лапы с перепонками действуют как весла; небольшие бугорки и полости-присоски на подошвах уменьшают опасность поскользнуться на льду, а плотная шерсть на подошвах защищает лапы от сильного холода и обеспечивает трение; уши меньше, чем у других медведей, и уменьшают потери тепла; веки действуют как солнечные очки; зубы острее, чем у других медведей, и больше подходят для полностью мясного рациона; увеличенный объем желудка позволяет голодному хищнику съесть сразу целого тюленя, кроме того белый медведь способен обходиться без пищи до девяти месяцев за счет переработки мочевины. Генетические данные свидетельствуют о том, что белый медведь (вид Ursus maritimus) отделился от североамериканских бурых медведей сравнительно недавно, всего около 150 тыс. лет назад, и очень быстро приспособился к своей новой среде обитания в Арктике. Этот вывод основан, в частности, на результатах анализа ДНК из челюсти ископаемого медведя, жившего 110-130 тыс. лет назад на Шпицбергене. Этот доисторический медведь по структуре своей ДНК оказался переходным между бурыми и белыми медведями (Lindqvist et al., 2009).
Эволюционное дерево
Филогенетическое дерево с указанием размера геномов. Классификация живых организмов представляет собой многоуровневую иерархическую структуру: организмы делятся на царства, царства делятся на типы, типы — на классы, классы — на отряды, и так далее. В результате такого ветвления получается филогенетическое дерево. Наличие единственной (естественной) классификации означает, что существует объективная закономерность в основе этой классификации. Именно такой результат можно ожидать при эволюционном происхождении животных от общего предка. Ветвление филогенетического дерева соответствует делению популяций в процессе видообразования. Несмотря на многочисленные разногласия между биологами по поводу отнесения тех или иных видов к конкретным группам (таксонам), эти противоречия имеют частный характер. Практика показывает, что биологические классификации, построенные на основе разных признаков (морфологических, эмбриологических, биохимических или генетических) в тенденции стремятся к одной и той же древовидной иерархической схеме — естественной классификации, отражающей последовательность расхождения эволюционных линий. Чем больше признаков учитывается в ходе классификации, тем выше сходство получаемых деревьев. Наличие естественной классификации было очевидно биологам еще в додарвиновские времена, и это изначально трактовалось как свидетельство иерархической организации замысла Творца. Однако в разнообразии других природных объектов, которые, в отличие от живых организмов, не происходят от общего предка, отсутствует единая древовидная иерархическая структура. Классификация таких объектов либо получается принципиально различной при использовании разных наборов признаков (например, минералы), либо имеет принципиально не «древесный» вид (например, химические элементы, звезды). Невозможно объективно построить иерархию элементарных частиц, химических элементов, планет Солнечной системы. Также не существует объективной иерархии таких сознательно созданных объектов, как книги в библиотеке, дома, мебель, машины и т. д.. Можно при желании объединять эти объекты в произвольные иерархии, но нет единственной объективной иерархии, принципиально лучшей, чем все остальные. Если бы была верна концепция «бараминов», естественная классификация и филогенетические деревья, получаемые на основе различных признаков и статистических методов, должны были бы иметь весьма характерный вид: изолированные пучки коротких ветвей (барамины) должны были бы отделяться друг от друга очень длинными (точнее, имеющими абсолютно произвольную длину) ветвями. Этого не наблюдается. Длины ветвей филогенетического дерева (отражающие, в первом приближении, степень различий по используемым признакам) в среднем примерно одинаковы между видами одного рода, родами одного семейства, семействами одного отряда и т.д. Это обстоятельство является дополнительным свидетельством в пользу того, что между процессами появления новых видов, родов, семейств, отрядов, классов, типов и т.д. не существует принципиальной разницы. Каждый новый род изначально появляется «всего лишь» как новый вид, и лишь позже, в ретроспективе, может «заслужить» в глазах биологов статус рода. Кроме того, если бы в идее «бараминов» было хоть какое-то рациональное зерно, этого не могли бы не заметить специалисты по систематике животных и растений еще задолго до Дарвина. Они бы обязательно увидели, что один из надвидовых таксономических рангов (род, отряд или семейство — антиэволюционисты обычно утверждают, что их «барамины» соответствуют каким-то из этих рангов) радикально отличается от всех остальных. Например, они бы увидели, что животный мир четко делится на семейства, которые поэтому являются особым, «естественным» уровнем классификации (и соответствуют «богосотворенным родам»), и гораздо менее четко делится на роды, подсемейства, надсемейства, отряды, подклассы и т.д., которые никаким «богосотворенным родам» не соответствуют. В действительности все ранги биологической классификации имеют примерно одинаковую степень как «четкости», так и «расплывчатости». По-настоящему «естественной» единицей классификации безоговорочно признается лишь вид (по критерию полного или почти полного отсутствия скрещиваний с другими видами в природе), но барамин по определению — это не вид, а нечто большее. Однако никакого надвидового ранга, более «естественного», чем остальные, в природе не существует. Существуют различные статистические методы для точной оценки того, насколько объекты с разными свойствами укладываются в ту или иную иерархию. Эти методы измеряют так называемый «филогенетический сигнал», позволяя отличить мнимые закономерности от объективных. Например, у любого «генеалогического древа» автомобилей будет низкий уровень филогенетического сигнала. У эволюционного дерева и у различных его частей, напротив, стабильно четкий филогенетический сигнал. Есть несколько источников данных, на основе которых можно делать выводы о степени родства между видами. Если существует единое эволюционное дерево, объединяющее все виды в объективную генеалогию, то все данные должны подтверждать эту генеалогию. Филогенетические деревья, построенные независимо, должны соответствовать друг другу. Наиболее простой, хотя и не очень точный способ построить такое дерево — сравнение строения организмов животных: чем меньше различий между видами, тем меньше поколений отделяют их от общего предка. Палеонтологическая летопись, наряду с другими подходами, позволяет установить родство между классами животных. Например, найденные останки пернатых динозавров свидетельствуют о родстве между рептилиями и птицами. Биогеография и эмбриология также дают информацию об эволюционной близости видов. Наиболее точный источник данных, недоступный во времена Дарвина — сравнительный анализ геномов различных организмов. Эволюционное дерево можно построить по каждому отдельно взятому гену, также исследователи рассматривают всевозможные некодирующие последовательности. Практика показывает, что чем больше генов включается в анализ, тем меньше остается в дереве статистически слабо обоснованных участков, и тем меньше различия между деревьями, построенными по разным наборам генов. Все эти источники информации дают одинаковую картину с точностью до погрешности используемых методов. Тот факт, что эволюционные деревья, построенные по разным данным, соответствуют друг другу, элементарно объясняется эволюционной теорией. Другие объяснения отсутствуют: например, нет никаких причин, почему организмы, сходные по строению, должны иметь сходные некодирующие последовательности в геноме (интроны в одних и тех же местах генов, остатки встроившихся ретровирусов в одних и тех же местах генома и т.д.), если они не произошли от одного предка. Примеры биологических исследований, связанных с построением эволюционных деревьев: 1) Новые данные позволили уточнить родословную животного царства 2) Невероятное разнообразие жуков получило эволюционное объяснение 3) Геном ланцетника помог раскрыть секрет эволюционного успеха позвоночных 4) Найдено «недостающее звено» между членистоногими и загадочными кембрийскими чудовищами аномалокарисами 5) Доказан скачкообразный характер эволюционного процесса Горизонтальный перенос генов, межвидовая гибридизация и симбиогенез порождают гориз
|
||||
|
|
Последнее изменение этой страницы: 2016-09-20; просмотров: 226; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.105.85 (0.015 с.) |