
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Ультраструктура химического синапса.Содержание книги Поиск на нашем сайте
Основными и наиболее изученными синаптическими образованиями играющими роль в деятельности нервной системы являются химические синапсы (Рис. 5).
Повышенная химическая чувствитель-ность постсинаптичес-кой мембраны является важным свойством си-напса, регулирующим его активность. Рис. 5. Структура химического синапса.
Активность синапса заключается в синаптической передаче - распространении возбуждения через синапс. Этот процесс является достаточно сложным и протекает в несколько стадий. 1. Синтез медиатора. Химические медиаторы, это вещества различной химической природы, среди которых различают моноамины (ацетилхолин, дофамин, норадреналин, серотонин и др.); аминокислоты (гамма-аминомасляная кислота - ГАМК, глутаминовая кислота, глицин и др.); нейропептиды (вещество Р, эндорфины, нейротензин, АКТГ, ангиотензин, вазопрессин, соматостатин и др.), поэтому их биосинтез разнообразен. Ацетилхолин, например, синтезируется из холина (продукт секретируемый печенью) и уксусной кислоты при участии ферментов нервной клетки; норадреналин синтезируется из аминокислот тирозина и фенилаланина. Медиаторы непрерывно синтезируются в нервной клетке и депонируются (складируются) в синаптических пузырьках нервных окончаний куда они поступают: во-первых из околоядерной области тела нейрона с помощью быстрого аксонального транспорта, во-вторых за счет синтеза медиатора, протекающего в самих синаптических окончаниях из продуктов его расщепления. Кроме того, медиатор может поступать в синаптическое окончание за счет обратного захвата его из синаптической щели в неизменном виде.
Следует отметить, что скорость биосинтеза медиатора невелика, и при больших нагрузках на синапс, может значительно отставать от скорости его разрушения. Работоспособность синапса при этом снижается. Именно эта причина является основной, при развитии всех видов утомления и время требуемое на отдых, это время требуемое на восстановление количества медиатора необходимого для полноценной работы синапса. 2. Секреция медиаторов.
Скорость экзоцитоза зависит от количества входящего кальция и активности метаболических процессов. Она увеличивается при введении лекарств содержащих кальций, АТФ, биогенных стимуляторов (алоэ, женьшень, боярышник), гормонов.
3. Взаимодействие медиатора с рецепторами постсинаптической мембраны.
Отметим, что электроуправляемых каналов на постсинаптической мембране нет, поэтому сама к генерации ПД она не способна. Возбуждающий постсинаптический потенциал (ВПСП), возникающий в результате поступления в клетку ионов Na+, электротонически распространяется за пределы постсинаптической мембраны и если его величина достаточно велика, деполяризует уже плазматическую мембрану до критического уровня. В мышечных клетках плазматическая мембрана сразу за пределами синапса имеет потенциал-чувствительные каналы, на ней относительно легко возникает полноценный потенциал действия. В нервных клетках возникновение возбуждения возможно только в аксоном холмике, поэтому активности одного из синапсов, которые, как правило, расположены далеко от него недостаточно для возникновения ПД. Нервная клетка воспринимает (суммирует) несколько синаптических сигналов и генерирует потенциал действия избирательно. В этом заключается интегративная функция нейрона.
Тормозной постсинаптический (ТПСП) потенциал возникает при усилении проницаемости для калия и хлора, он гиперполяризует постсинаптическую мембрану, препятствуя электротоническому распространению возбуждения по мембране нервной клетки. Процесс торможения имеет место только на межнейронных синапсах. Принципы работы возбуждающего и тормозного синапсов сходны. Разница заключается в том с какими специфическими каналами связаны рецепторы расположенные на постсинаптической мембране, от чего и зависит физиологическая реакция.
4. Инактивация медиатора
Продукты расщепления либо поступают в кровь и циркулируют как его предшественники, либо за счет пиноцитоза осуществляется их «обратное всасывание» в синаптическое окончание, где происходит их восстановление и упаковка в синаптические пузырьки и медиатор может использоваться повторно.
Свойства синапсов: · односторонность проведения возбуждения через синапс (всегда от пре- к постсинаптической мембране обусловлена наличием рецепторов чувствительных к медиатору только на постсинаптической мембране; · синаптическая задержка - замедление скорости распространения возбуждения в синапсе в связи с длительностью процессов выделения медиатора из пресинаптического окончания, диффузии его по синаптической щели и процесса взаимодействия его с постсинаптической мембраной. Синаптическая задержка в химических синапсах обычно равна 0,2 - 0,5 мс. · низкая лабильность - обусловлена временем распространения предыдущего импульса и наличием у него периода абсолютной рефрактерности; · повышенная утомляемость - обусловлена отставанием скорости синтеза медиатора от скорости его распада при возбуждении; · трансформация ритма возбуждения, обусловлена тем, что не все приходящие к синаптическому окончанию аксона потенциалы действия способны вызвать критическую деполяризацию постсинаптической мембраны;
· высокая чувствительность к лекарствам и ядам обусловлена высокой специфичностью хеморецепторов постсинаптической мембраны; · низкий фактор безопасности (надежности), обусловлен тем, что хеморецепторы чувствительны к большому количеству веществ, способных инициировать или изменять активность синапса.
АДРЕНЕРГИЧЕCКИЕ СИНАПСЫ Адренергические нейроны расположены в ЦНС (голубое пятно среднего мозга, мост, продолговатый мозг) и в симпатических ганглиях. Периферические адренергические синапсы образованы варикозными утолщениями разветвлений постганглионарных симпатических волокон. Медиатор адренергических синапсов - норадреналин. Его предшественник в биосинтезе дофамин выполняет медиаторную функцию в дофаминергических синапсах. Адреналин представляет собой гормон мозгового слоя надпочечников. Все три вещества относятся к группе катехоламинов, так как содержат гидроксилы в 3-м и 4-м положениях ароматического кольца. Синаптические пузырьки в адренергических синапсах имеют под электронным микроскопом гранулярное строение и поэтому получили название гранулы. В гранулах норадреналин депонирован в связи с АТФ и белком хромогранином. В составе гранул обнаружены также ферменты и модулирующие нейропептиды (энкефапины, нейропептид Y). Норадреналин синтезируется из аминокислоты тирозина. Превращение фенилаланина в тирозин является неспецифическим процессом и происходит в печени. Обе аминокислоты в большом количестве содержатся в твороге, сыре, шоколаде, бобовых. ТИРОЗИН
(тирозин-3-монооксигеназа)
ДОФА
L- аминокиспот Кофактор - пиридоксальфосфат
Дофамин-β-гидроксилаза Кофактор – аскорбат
Фенилэтаноламин-N-метилтрансфераза АДРЕНАЛИН
После диссоциации комплексов норадреналин - адренорецептор медиатор инактивируется при участии ряда механизмов: • Нейрональный захват (захват-1) - активный транспорт вначале через пресинаптическую мембрану (сопряжен с выходом ионов натрия), а затем через мембрану гранул под влиянием АТФ-зависимой протонной транслоказы (при входе в гранулы одной молекулы норадреналина в цитоплазму выходят 2 протона);
• Экстранейрональный захват (захват-2) нейроглией, фибробластами, миокардом, эндотелием и гладкими мышцами сосудов; • Инактивация ферментами. 70-80% норадреналина участвует в нейрональном захвате, по 10% подвергается экстранейрональному захвату и ферментативному расщеплению. Необходимость нейронального захвата диктуется дефицитом субстратов и большой потребностью в энергии для синтеза норадреналина из тирозина. Для сохранения адреналина основное значение имеет экстранейрональный захват. Ферменты инактивации катехоламинов - моноаминоксидаза (МАО) и катехол – О -метилтрансфераза (КОМТ). МАО, локализованная на внешней мембране митохондрий и в гранулах, осуществляет окислительное дезаминирование катехоламинов с образованием биогенных альдегидов. Затем альдегиды окисляются НАД-зависимой альдегиддегид-рогеназой в кислоты или восстанавливаются альдегидредуктазой в гликоли. Цитоплазматический фермент КОМТ катализирует присоединение метильной группы к гидроксилу в 3-м положении ароматического кольца (только при наличии гидроксила в 4-м положении). Донатором метильных групп служит S-аденозилметионин. Метилированные продукты в 200-2000 раз (по разным тестам) менее активны, чем норадреналин и адреналин. Адренорецепторы Адренорецепторы локализованы на постсинаптической, пресинаптической мембранах и в органах, не получающих адренергическую иннервацию. Постсинаптические адренорецепторы имеют индексы 1 или 2, пресинаптические и внесинаптические адренорецепторы обозначаются индексом 2. Внесинаптические адренорецепторы возбуждаются циркулирующимив крови норадреналином и адреналином. Постсинаптические α1,-адренорецепторы (А, В, О) регулируют функцию мембранных фосфолипаз и проницаемость кальциевых каналов L -типа. В гладких мышцах ионы кальция активируют кальмодулин - зависимую киназу легких цепей миозина, что необходимо для образования актомиозина и сокращения. Только в желудке и кишечнике α1-адренорецепторы, открывая кальцийзависимые калиевые каналы, вызывают гиперполяризацию сарколеммы и расслабление гладких мышц. Эффекты возбуждения α1-адренорецепторов: · Сокращение радиальной мышцы радужки с расширением зрачков (мидриаз; греч. amydros - темный, неясный); · Сужение сосудов кожи, слизистых оболочек, органов пищеварения, почек и головного мозга; · Повышение АД; · Сокращение капсулы селезенки с выбросом депонированной крови; · Сокращение сфинктеров пищеварительного тракта и мочевого пузыря; · Уменьшение моторики и тонуса желудка и кишечника. α 2 - Адренорецепторы (А, В, С) снижают активность аденилатциклазы. Постсинаптические α2 - адренорецепторы суживают сосуды кожи и слизистых оболочек, тормозят моторику желудка и кишечника, уменьшают секрецию кишечного сока. Пресинаптичесие α2 - адренорецепторы по принципу отрицательной обратной связи снижают выделение норадреналина из адренергических окончаний при избытке медиатора в синаптической щели (увеличивают калиевую проводимость мембран, блокируют кальциевые каналы L - и N -типов).
Внесинаптические α2-адренорецепторы вызывают спазм сосудов, подавляют секрецию инсулина и повышают агрегацию тромбоцитов. β-Адренорецепторы, активируя аденилатциклазу, повышают синтез цАМФ (β1,-адренорецепторы также открывают кальциевые каналы в миокарде). Для постсинаптических β1-адренорецепторов характерны следующие эффекты: · Возбуждение сердца - тахикардия, ускорение проведения импульсов по проводящей системе, усиление сокращений миокарда, рост потребности в кислороде, неадекватный выполняемой работе; · Ослабление моторики кишечника; · Секреция ренина; · цАМФ-зависимый липолиз в жировых депо. Постсинаптические и внесинаптические β2-адренорецепторы расслабляют гладкие мышцы и вызывают гипергликемию: • Расширяют сосуды сердца, легких и скелетных мышц; • Снижают АД; • Расширяют бронхи и уменьшают секреторную функцию бронхиальных желез; •Тормозят моторику желудка и кишечника; • Расслабляют желчный пузырь, мочевой пузырь, беременную и небеременную матку; • Усиливают цАМФ-зависимые гликогенолиз и гликонеогенез в печени, гликогенолиз в скелетных мышцах; • Повышают секрецию инсулина. Пресинаптические β2-адренорецепторы осуществляют положительную обратную связь, стимулируя выделение норадреналина при его дефиците в синаптической щели. В сосудах и внутренних органах расположены α- и β-адренорецепторы различных типов, например, в сосудах легких обнаружено 30% β1-адренорецепторов и 70% β2-адренорецепторов.
|
|||||||||
|
|
Последнее изменение этой страницы: 2016-08-26; просмотров: 403; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.222.212.80 (0.012 с.) |

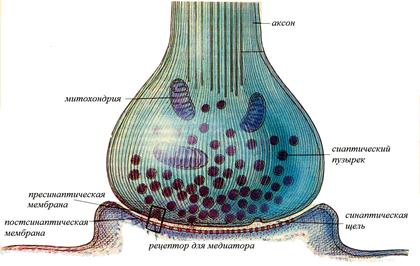 Их пресинаптическое окончание образует характерное утолщение, покрытое пресинаптической мембраной. Между нервным окончанием и эффекторной клеткой имеется пространство называемое синапти-ческой щелью, которая отделяет нервное окончание от мембраны эффекторной клетки. Часть мембраны эффекторной клетки лежащая за синаптической щелью называется постсинаптической мембраной. Эта мембрана имеет белковые хеморецепторы, которые связаны с хемо-управляемыми ионными каналами. Связывание рецепторов с биологи-чески активными веще-ствами (медиаторами, гормонами, лекарст-венными препаратами), приводит к открытию канала, движению ионов через него и изменению заряда мембраны.
Их пресинаптическое окончание образует характерное утолщение, покрытое пресинаптической мембраной. Между нервным окончанием и эффекторной клеткой имеется пространство называемое синапти-ческой щелью, которая отделяет нервное окончание от мембраны эффекторной клетки. Часть мембраны эффекторной клетки лежащая за синаптической щелью называется постсинаптической мембраной. Эта мембрана имеет белковые хеморецепторы, которые связаны с хемо-управляемыми ионными каналами. Связывание рецепторов с биологи-чески активными веще-ствами (медиаторами, гормонами, лекарст-венными препаратами), приводит к открытию канала, движению ионов через него и изменению заряда мембраны. Высвобождение медиатора из синаптических пузырьков осуществляется дискретно, квантами (порциями). В состоянии покоя оно незначительно, и не вызывает никакого эффекта, но под влиянием нервного импульса, приходящего по аксону, резко усиливается. Возбуждение, деполяризует мембрану синаптического окончания, что приводит к активации имеющихся на ней Са2+-каналов и входу ионов кальция в синаптическое окончание. Пузырьки находящиеся у пресинаптической мембраны, под действием Са2+ разрушаются (по механизму экзоцитоза) и медиатор попадает в синаптическую щель. Медиатор выделившийся из синаптических пузырьков пресинаптического окончания диффундирует в синаптической щели и достигает постсинаптической мембраны.
Высвобождение медиатора из синаптических пузырьков осуществляется дискретно, квантами (порциями). В состоянии покоя оно незначительно, и не вызывает никакого эффекта, но под влиянием нервного импульса, приходящего по аксону, резко усиливается. Возбуждение, деполяризует мембрану синаптического окончания, что приводит к активации имеющихся на ней Са2+-каналов и входу ионов кальция в синаптическое окончание. Пузырьки находящиеся у пресинаптической мембраны, под действием Са2+ разрушаются (по механизму экзоцитоза) и медиатор попадает в синаптическую щель. Медиатор выделившийся из синаптических пузырьков пресинаптического окончания диффундирует в синаптической щели и достигает постсинаптической мембраны.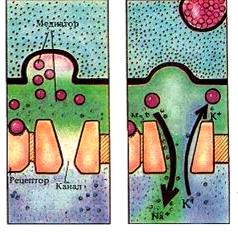 Медиатор достигший постсинаптической мембраны действует на ее хеморецепторы, связанные с различными хемо-управляемыми ионными каналами, вследствие чего изменяется проницаемость постсинаптической мембраны для различных ионов (Na+, K+, Cl-). В зависимости от иона и направления его движения постсинаптическая мембрана деполяризуется или гиперполяризуется и на ней возникает возбуждающий или тормозной потенциал.
Медиатор достигший постсинаптической мембраны действует на ее хеморецепторы, связанные с различными хемо-управляемыми ионными каналами, вследствие чего изменяется проницаемость постсинаптической мембраны для различных ионов (Na+, K+, Cl-). В зависимости от иона и направления его движения постсинаптическая мембрана деполяризуется или гиперполяризуется и на ней возникает возбуждающий или тормозной потенциал. Инактивация медиатора - последняя стадия синаптической передачи, обеспечивающая ее прекращение. Она необходима для реполяризации постсинаптической мембраны и восстановления исходного уровня ее мембранного потенциала, что обеспечивает возможность восприятия следующего сигнала. Наиболее распространенным является гидролитическое расщепление медиатора с помощью соответствующих ферментов, постоянно находящихся в синаптической щели. Для ацетилхолина, например, таким ферментом является холинэстераза, для норадреналина - моноаминооксидаза. Эти ферменты начинают разрушать медиатор на исходные компоненты, сразу, как только он попадает в синаптическую щель, и скорость разрушения настолько велика, что при небольших количествах медиатора, он разрушается еще до того, как достигает постсинаптической мембраны.
Инактивация медиатора - последняя стадия синаптической передачи, обеспечивающая ее прекращение. Она необходима для реполяризации постсинаптической мембраны и восстановления исходного уровня ее мембранного потенциала, что обеспечивает возможность восприятия следующего сигнала. Наиболее распространенным является гидролитическое расщепление медиатора с помощью соответствующих ферментов, постоянно находящихся в синаптической щели. Для ацетилхолина, например, таким ферментом является холинэстераза, для норадреналина - моноаминооксидаза. Эти ферменты начинают разрушать медиатор на исходные компоненты, сразу, как только он попадает в синаптическую щель, и скорость разрушения настолько велика, что при небольших количествах медиатора, он разрушается еще до того, как достигает постсинаптической мембраны. Тирозингидроксилаза
Тирозингидроксилаза Кофактор - тетрагидроптерин
Кофактор - тетрагидроптерин Декарбоксилаза ароматических
Декарбоксилаза ароматических ДОФАМИН
ДОФАМИН НОРАДРЕНАЛИН
НОРАДРЕНАЛИН


