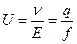Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Виникнення клітинної організаціїСодержание книги
Поиск на нашем сайте БІОФІЗИКА КЛІТИНИ КЛІТИНА Клітина – найменша структурна одиниця всього живого, здійснює обмін речовин та енергії, росте, розмножується, і передає по спадковості свої ознаки, реагує на подразнення, має властивість рухатися. Здатність клітин до використання і перетворення енергії має надзвичайно важливе значення, оскільки клітина зберігає постійність внутрішнього середовища і цілісність її структури. ІСТОРІЯ БІОФІЗИКИ Поштовхом розвитку біофізики клітини була клітинна теорія Шванна. Роботи вченого відносяться до різноманітних галузей біології: · Дія кисню на розвиток птахів з яєць; · Процес гниття; · Бродіння з участю дріжджів; · Відкрив пепсин у 1836 р. · Досліджував клітинну будову хряща личинок земноводних і хорди під мікроскопом;
Сучасний стан КЛІТИННОЇ ТЕОРІЇ: 1. Клітина – основна найменша структурно-функціональна та генетична одиниця живих організмів. 2. Клітини всіх організмів принципово подібні за будовою, хімічним складом і важливими проявами процесів життєдіяльності. 3. Кожна нова клітина утворюється в результаті поділу материнської клітини. 4. Клітини багатоклітинних організмів спеціалізовані за виконанням різноманітних функцій та утворюють тканини. 5. В основі розмноження, росту, розвитку і регенерації організмів лежить поділ, ріст і диференціювання клітин. 6. Клітина – відкрита система, існує тісний зв'язок з оточуючим середовищем, через неї проходить потік речовини, енергії та інформації. Виникнення клітинної організації 1 етап: прокаріотичні гетеротрофні анаеробні клітини (3,5 млрд.р.). 2 етап: прокаріотичні автотрофні анаеробні клітини. 3 етап: прокаріотичні автотрофні аеробні клітини. 4 етап: еукаріотичні авто- і гетеротрофні аеробні клітини(1,5 млрд. р.). Гіпотези походження еукаріот: СИМБІОЗНА: симбіоти клітини-господаря – анаеробний прокаріот;
ІНВАГІЦІЙНА: вихідна клітина – аеробний прокаріот. КЛІТИННІ МЕМБРАНИ Клітинні мембрани – високоорганізовані структури клітини, які виконують ряд функцій: 1. представляють собою напівпроникні бар’єри – захисна функція для клітин на внутрішньоклітинних органел; 2. здійснюють вибірковий транспорт різноманітних речовин всередину і назовні клітини; 3. передача інформації з допомогою гормонів, медіаторів, нервових імпульсів; 4. перетворення енергії (синтез АТФ здійснюється на внутрішніх мембранах мітохондрій за рахунок енергії трансмембранного градієнта концентрації протонів); 5. процеси молекулярного впізнавання проходять на мембранах клітин, де розміщені рецептори гормонів, молекули імунної системи; 6. ферментативна діяльність пов’язана з координацією всіх біохімічних реакцій, що протікають в клітині. Термін “ мембрани ”, як невидима плівка, що оточує клітину, вперше використано ботаніком фон Молем і незалежно К. фон Негелі (1817-1891) в 1855 році для пояснення явища плазмолізу.
Склад клітинних мембран залежить від їхнього типу та функцій, однак їхній склад є майже універсальним: ü Ліпіди (50%); ü Білки (40%); ü Вуглеводи (10%). Вчені Е. Гортер і Ф. Грендель виміряли площу моношару, сформованого ліпідами, які екстрагували ацетоном з еритроцитів, і порівняли з площею поверхні цих клітин. Виявили, що площа моношару вдвічі перевищує площу поверхні клітин. Таким чином дійшли висновку, що ліпіди у клітинній мембрані формують бімолекулярний шар (ліпідний бішар) – така думка є загальновизнаною. МОДЕЛІ МЕМБРАН Двошарова модель мембрани, включає 2 шари ліпідів, Бутербродна модель мембрани Даніелі та Давсона (1935), включає білки, та враховує проникність мембрани для іонів та аніонів. Двошарова модель мембрани Робертсона (1958), включає шар ліпідів всередині шару білків. «РІДИННО-МОЗАЇЧНА» МОДЕЛЬ МЕМБРАН Нікольсона і Сінгера. У 1972 році С. Сінгер та
МЕМБРАННІ ЛІПІДИ Мембранні ліпіди – це амфифільні (амфіпатичні, amphipathic) сполуки, тому що вони складаються з полярної гідрофільної групи (“головки”) й більш протяжної гідрофобної групи (“хвоста”). СТРУКТУРА МЕМБРАН Зовнішня цитоплазматична мембрана, як тваринних, так і рослинних клітин, має трьохшарову (триламінарну) структуру (товщина 5-10 нм): • 1) надмембранний компонент - глікокалікс; • 2) плазмалема; • 3) підмембранний шар – опорно-скоротливий комплекс - цитоскелет.
МЕМБРАННІ ЛІПІДИ На поверхні води ліпіди формують моношар, в якому полярні групи молекул контактують з водою, а неполярні ланцюги орієнтовані всередину під прямим кутом до моношару. Розрізняють три класи ліпідів, котрі входять до клітинних мембран: ü фосфоліпіди; ü сфінголіпіди; ü холестерол.
Бішар ліпідів стабілізується завдяки гідрофобним зв'язкам або Ван-дер-Ваальсовим взаємодіям. СТРУКТУРА ФОСФОЛІПІДІВ ФОСФОЛІПІДИ Фосфоліпіди – їх загалом називають фосфоацилгліцеролами, до складу входять 2 залишки жирних кислот, залишок фосфорної кислоти, яка має складноетерний зв'язок з іншими молекулами. Фосфоліпіди є не лише структурними компонентами клітин, але також можуть виконувати важливі біохімічні та фізіологічні функції у клітинах. Наприклад, арахідонова кислота є попередником важливих фізіологічних посередників – простогландинів, тромбоксанів. Фосфоліпіди – як видно з назви складаються з залишків фосфорної кислоти, і є похідними трьохатомного спирту гліцерину або складного спирту сфінгозину. ФЕА – міститься в тканинах тварин і в меншій кількості у рослин (15-30%), однак це основний фосфоліпід клітин бактерій. ФС – мінорний фосфоліпід, але входить до мембран про- і еукаріот, найбільше його у клітинах мозку (15%), печінки, серця. ДФГ – міститься у внутрішній мембрані мітохондрій, хлоропластів, рідко в бактеріях. ФХ – основний компонент мембранних фосфоліпідів (50%), в бактеріальних клітинах його немає. ФОСФОЛІПІДИ
ГЛІЦЕРОФОСФОЛІПІДИ МЕМБРАН Полярна «головка» ГЛІЦЕРОЛ ФОСФАТ ХОЛІН Гідрофобний «хвіст» ЖИРНА КИСЛОТА Жирні кислоти входять до складу фосфоліпідів, тригліцеролів і сфінголіпідів. гліцерофосфоліпіди сфінголіпіди триацилгліцероли СФІНГОЛІПІДИ Сфінгомієліни – найпростіші й найпоширеніші з сфінголіпідів, містять залишок фосфору, тоді як інші два підкласи не містять його, несуть електричний заряд; містяться в більшості мембран тваринних клітин, оосбливо багато у оболонках нервових волокон. Сфінголіпіди – мають також полярну голівку, та 2 неполярні хвости, один з яких представлений залишком дволанцюгового аміноспирту сфінгозину, однак не містять гліцеролу. Існують три підкласи сфінголіпідів: 1) сфінгомієліни, 2) цероброзиди, 3) гангліозиди. Гангліозиди – найбільш складні ліпіди мембран, їхні голівки утворені кількома залишками цукрів. Несуть негативний заряд, оскільки містятб один або декілька залишків сіалової кислоти. Вперше їх виділили з гангліїв, найбільш багатий на гангліозиди мозок (5-10%), нирки, печінки. Гангліозиди локалізуються в плазматичних мембранах і відповідають за контактне гальмування, адгезію, електрофоретичну рухливість, приймають участь у процесах вибіркового транспорту і володіють імунною активністю. Цероброзиди – не мають електричного заряду, оскільки їхні полярні голівки утворені електронейтральними групами, вони належать до гліколіпідів, оскільки несуть у своєму складі залишки цукрів. СТЕРОЛИ Холестерин зустрічається в плазматичних мембранах тварин, в лізосомах, ендосомах, апараті Гольджі; у рослин поширені сито - і стигмастероли, в мембранах дріжджів та прокаріот ергостерол. ПОЛОЖЕННЯ ХОЛЕСТЕРОЛА В ЛІПІДНОМУ БІШАРІ «Полярні Головки» Ділянка Холестерола, який надає жорсткості мембрані Ділянка гідрофобного хвоста, який володіє меншою вязкістю ДИНАМІКА ЛІПІДІВ У МЕМБРАНІ Найважливішою фізичною властивістю дволанцюгових амфіфільних молекул є їхня схильність утворювати подвійні (бішари) у водному середовищі. Бішари формують окремі пухирці (везикули) або пластинчасті (ламелярні) структури. МЕТОДИ ДОСЛІДЖЕННЯ Для тонкого вивчення структури клітинних мембран застосовують ЕПР (електронний парамагнітний резонанс), ЯМР (ядерний магнітний резонанс) та рентгеноструктурний аналіз. Дифракція рентгенівських променів – найефективніший сучасний метод дослідження структури великих молекул. Дані про структуру бішарів було отримано за допомогою ЕПР, сигнал ЕПР дає тільки неспарений електрон, і у звичайного бішару він відсутній, тому вводять у бішар спінові мітки (нітроксильні радикали). Спектр бішару суттєво змінюється зі зміною його структури. Продовження · Особливості та характеристика мембранних білків; · Асиметрія мембран; · Динаміка мембранних білків; · Методи дослідження мембранних білків; · Функції мембранних білків; · Мембранні вуглеводи - глікокалікс; · Цитоскелет; · Роль мембран у старінні клітин; · Дія стресових чинників на мембрани. МЕМБРАННІ БІЛКИ Мембрани різняться між собою за вмістом білка. Наприклад, у мієліновій оболонці невелика кількість білка, приблизно 18% від загальної маси мембрани.
В інших клітинах загальна маса білка становить до 50%, а в мембранах мітохондрій та хлоропластах до 75%. Мембранні білки забезпечують специфічні властивості плазматичної мембрани. Відносно розміщення у ліпідному шарі мембранні білки поділяють на: Інтегральні білки; Напівінтегральні білки; Периферичні білки. ІНТЕГРАЛЬНІ БІЛКИ Ú Важко виділити; Ú Підтримують асоціацію з ліпідами; Ú Містять значну кількість гідрофобних АМ; Ú Асиметрично орієнтовані амфіфіли; Ú Трансмембранні білки. Пронизують подвійний бішар ліпідів, формують канал, з гідрофобною порою, крім гідрофільних ділянок, містять й гідрофобні. Ú Інтегральна частина – неполярні АК • гідрофобна • якір білка всередині мембрани Ú Частина над поверхнею мембрани – полярні АК • гідрофільна • поширена у позаклітинній рідині & цитозолі Тобто крім електростатичних взаємодій, з фрагментами мембран формують гідрофобні взаємодії. Представниками інтегральних білків є глікофорин А (глікопротеїн), бактеріородопсин, порини бактерій. ПЕРЕФЕРИЧНІ БІЛКИ Ú Легко виділити: – Висока концентрація іонів – Зміни pH середовища Ú Вільно прикріплені до ліпідів; Ú Розчинні у воді (e.g. cytochrome c); Ú Нормальний амінокислотний склад. Розміщені на периферії подвійного бішару ліпідів і не формують сильних зв'язків з його компонентами. Зв'язані з полярними головками ліпідів або/чи приєднані до цитоплазматичних ділянок інтегральних білків Зв'язані з допомогою електростатичних зв'язків. Легко висолюються або детергуються ЕДТА. Переважно це глобулярні білки, однак фібрилярні білки, котрі входять до склад цитоскелету, також належать до переферичних. АСИМЕТРІЯ МЕМБРАН Трансмембранна асиметрія мембрани свідчить про те, що різні половини бішару має різний склад. Встановлено, що інтегральні мембранні білки вмонтовані в мембрану АСИМЕТРИЧНО і ця асиметрія є стабільною. Існують дані про латеральну гетерогенність біологічних мембран, це можуть бути досить значні спеціалізовані ділянки мембрани.
ДИНАМІКА МЕМБРАН Багато мембранних білків здатні до динаміки. Однак рухи білків частково є обмежені через взаємодію з цитоскелетом та з іншими мембранними білками. Рухливість мембранних білків показали на експериментальному змішуванні мембранних білків клітини миші та людини, з додаванням флоресцентної мітки. ДІАЛІЗ Використовують для виділення низькомолекулярних домішок або заміни складу середовища ХРОМАТОГРАФІЯ Гель-проникаюча хроматографія дозволяє розділяти білки за розміром і формою молекул. Поділ проводять у хроматографічних колонках заповнених сферичними частинками набухлого гелю із полімерних матеріалів. Білкові молекули, які не здатні проникати в гранули гелю, будуть переміщатися з високою швидкістю. Середні та невеликі білки будуть утримуватися гранулами гелю Електрофорез у поліакриламідному гелі в присутності додецилсульфату натрію Метод базується на властивості заряджених часток (молекул) переміщатися під дією електричного поля Електрофорез проводять у тонкому шарі поліакриламіду МЕТОД ЕЛЕКТРОФОРЕЗУ Електрофорез. Електрофорезом називається рух частинок в рідині під дією електричного поля. На макромолекулу із сумарним зарядом Q в електричному полі напруженістю Е діє електрична сила Fе: а – комплекс поліпептидного ланцюга (1) з молекулами ДСН (2);
U=b-a·lgM Кожна молекула ДСН має один негативний заряд, звідси загальна густина заряду приблизно однакова для різних білкових поліпептидних ланцюгів. Отже, поверхнева шуба з ДСН усуває зарядові відмінності нативних білків. Після денатурації поліпептид має вигляд витягнутого циліндра довжиною 1,8 нм. Відношення V до напруженості електричного поля Е називається електрофоретичною рухливістю U (м2· с-1·В-1):
де η – в'язкість середовища; r – радіус макромолекули.
Найкращим методом, за яким визначається молекулярна маса й субодиничний склад макромолекул, є електрофорез у поліакриламідному гелі у присутності додецилсульфату натрію (ДСН). Експериментально встановлено, що за певних умов електрофоретична рухомість залежить від молекулярної маси й на цю залежність не впливає загальний заряд макромолекули. До цього білок обробляють 1% розчином ДСН, яки є детергентом, що денатурує білки, крім чого додають β-меркаптоетанол для розриву дисульфідних зв'язків (1,4 кг ДСН на 1 кг білка). Одночасно проводять електрофорез досліджуваного білка та маркерних білків з відомою молекулярною масою й графічно зображають цю залежність. ІНШІ МЕТОДИ ДОСЛІДЖЕННЯ Метод радіаційної інактивації використовують для визначення розміру мішені, суть методу полягає у визначенні долі білкових молекул, які отримали пошкодження при опроміненні. Електрофізіологічні методи; Мікроскопія: Ú Заморожування-сколювання (вугілля або Pt); Ú Електронна та електронна скануюча; Ú Флуоресцентна. Також застосовують методи кругового дихроїзму, електронної мікроскопії, інфрачервону та раманівську спектроскопію, ЯМР. ФУНКЦІЇ МЕМБРАННИХ БІЛКІВ · Транспорт. (ліворуч) Білки, що пронизують мембрану наскрізь, формують гідрофільні канали, які селективні до певного виду іонів. (праворуч) Інші транспортні білки, в якості помп, використовуючи субстрат з цитоплазми клітини (наприклад, АТР), як джерело енергії, переносять молекули через мембрану. · Ферментативна активність. Білки, які вмонтовані в мембрану володіють ферментативною активністю, і включені в ланцюг метаболізму. · Передача сигналів. Мембранні білки на своїй поверхні містять спеціалізовані ділянки (сайти) до хімічних месенджерів, наприклад, гормонів. Зовнішній месенджер (гормон) викликає конформаційні зміни білка зв'язуючись із спеціалізованою ділянкою, що реалізує передачі сигналу (інформації) всередину клітини. · Клітинне впізнавання. Деякі глікопротеїди використовуються в якості якорів для клітинного впізнавання. · Міжклітинна взаємодія. Мембранні білки, двох контактуючих клітин формують високоспеціалізовані структури – міжклітинні контакти. · Взаємодія з цитоскелетом та матриксом. Мікрофіламенти та інші елементи цитоскелету можуть звязуватися з мембранними білками на своїй поверхні та стабілізувати локалізацію білків. МЕМБРАННІ ВУГЛЕВОДИ Ú Переважно олігосахариди; Ú Різноманітний склад моносахаридів; Ú Багато сіалової кислоти; Ú Гліколіпіди; Ú Глікопротеїни Вуглеводи в мембрані становлять приблизно від
Приклади, гліколіпідів та глікопротеїдів у плазматичній мембрані, які формують глікокалікс. Ú Відіграють ключову роль в cell-cell впізнаванні – Здатність клітин впізнавати “потрібну” клітину • antigens – Важливі для тканинного & – Основа для відторгнення клітин імунною системою ЦИТОСКЕЛЕТ Ú Інфраструктура клітини; Ú Підтримання форми клітини – Механічна підтримка – Підтримка органел Ú Рух – Поділ клітини ФУНКЦІЇ ЦИТОСКЕЛЕТУ v Задіяний у розходженні хромосом під час мітозу і подальшого поділу клітин; v Керує і направляє внутрішньоклітинний транспорт між органелами, переносячи матеріал від однієї частини клітини до іншої; v Підтримує клітинну мембрану; v Дозволяє деяким клітинам (сперматозоїди) плавати, а іншим (фібробласти і лейкоцити) переміщатися; v Відповідає за скорочення м’язових клітин; v Відповідає за ріст аксону та дендритів; v Контролює ріст клітинної стінки рослинних клітин; v Відповідає за різноманітність форми еукаріотичних клітин. ЕЛЕМЕНТИ ЦИТОСКЕЛЕТУ Ú Мікрофіламенти – Взаємодіють з мембранними білками – Активний рух – Основний елемент м'язів Ú Мікротрубочки – Клітинний скелет – Веретено волокна, війок джгутиків Ú Мікроворсинки – Захищають поверхню клітин – Збільшують поверхню
Актинові філаменти визначають форму поверхні клітини та є необхідними для переміщення усієї клітини. Але самі по собі ці філаменти неефективні. Їхня корисність для клітини залежить від цілого ряду допоміжних білків, які зв’язують ці філаменти з іншими компонентами клітини, як і з самими собою. Проміжні філаменти забезпечують механічну стійкість і опір проти зовнішнього тиску. Мікротрубочки визначають положення мембранних органел і прямий внутрішньоклітинний транспорт. СТРЕС Ú Стабільність клітинних мембран розглядають як інтегральний фактор стійкості до несприятливих умов середовища. Ú Мембрани першими піддаються дії стресових факторів. Вони представляють собою мішені первинного впливу! Ú Підвищується проникність мембрани, проходить деполяризація мембранного потенціалу плазмалеми, рН зміщується у кислу сторону. Зростає активність Н+-помпи в плазмалемі та тонопласті, посилюється збирання мікрофіламентів і цитоскелету, наслідком чого є зростання в'язкості цитоплазми. Збільшується ПОЛ та затрати АТФ на підтримку структури та обміну речовин. Ú Зростає швидкість процесів гідролізу і гальмується синтез білка. Ú Структурні перебудови в мембранах в значній мірі стосуються ліпідів, а саме можуть проходити зсуви різноманітних радикалів жирних кислот, змінюється степінь їх ненасиченості. Ú Структурні зміни в мембранах під впливом несприятливих факторів впливу стосуються і вивільнення з зв'язаного стану іонів Ca2+, який утворює мостики між карбоксильними групами білків та полярними головками фосфоліпідів і значно зростає його концентрація в цитоплазмі, такого короткочасного підвищення досить щоб запустити специфічні мембранні канали і транспортні системи. Ú Причини зростання проникності мембрани різноманітні. Їх повязують із зміною співвідношення Н+/Ca2+ в клітині.
Ú Стабілізатором клітинних мемран є іони Ca2+. В присутності кальцію проходить збільшення електричного опору мембрани. Він впливає на проникність мембрани для інших іонів, здійснює регуляцію траспорту води. Ú Таким чином, фундаментальну роль в стійкості клітин (особливо рослин) до несприятливих факторів відіграють клітинні мембрани, иякі інтегрують дію різноманітних факторів. СИНТЕЗ ТА ЗБИРАННЯ МЕМБРАН Ú Мембранні білки та ліпіди синтезуються та складаються в ендомембранних системах: ЕПР (гранулярний) та Комплексі Гольджі Ú У гладенькому ЕПР відсутні рибосоми – він відповідає за синтез ліпідів.
БІОФІЗИКА КЛІТИНИ КЛІТИНА Клітина – найменша структурна одиниця всього живого, здійснює обмін речовин та енергії, росте, розмножується, і передає по спадковості свої ознаки, реагує на подразнення, має властивість рухатися. Здатність клітин до використання і перетворення енергії має надзвичайно важливе значення, оскільки клітина зберігає постійність внутрішнього середовища і цілісність її структури. ІСТОРІЯ БІОФІЗИКИ Поштовхом розвитку біофізики клітини була клітинна теорія Шванна. Роботи вченого відносяться до різноманітних галузей біології: · Дія кисню на розвиток птахів з яєць; · Процес гниття; · Бродіння з участю дріжджів; · Відкрив пепсин у 1836 р. · Досліджував клітинну будову хряща личинок земноводних і хорди під мікроскопом;
Сучасний стан КЛІТИННОЇ ТЕОРІЇ: 1. Клітина – основна найменша структурно-функціональна та генетична одиниця живих організмів. 2. Клітини всіх організмів принципово подібні за будовою, хімічним складом і важливими проявами процесів життєдіяльності. 3. Кожна нова клітина утворюється в результаті поділу материнської клітини. 4. Клітини багатоклітинних організмів спеціалізовані за виконанням різноманітних функцій та утворюють тканини. 5. В основі розмноження, росту, розвитку і регенерації організмів лежить поділ, ріст і диференціювання клітин. 6. Клітина – відкрита система, існує тісний зв'язок з оточуючим середовищем, через неї проходить потік речовини, енергії та інформації. Виникнення клітинної організації 1 етап: прокаріотичні гетеротрофні анаеробні клітини (3,5 млрд.р.). 2 етап: прокаріотичні автотрофні анаеробні клітини. 3 етап: прокаріотичні автотрофні аеробні клітини. 4 етап: еукаріотичні авто- і гетеротрофні аеробні клітини(1,5 млрд. р.).
|
|||||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 529; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.216 (0.017 с.) |

 б – експериментально встановлено лінійний зв'язок між електрофоретичною рухливістю U й логарифмом молекулярної маси М (контрольні білки);
б – експериментально встановлено лінійний зв'язок між електрофоретичною рухливістю U й логарифмом молекулярної маси М (контрольні білки);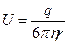 Під дією цієї сили відбувається прискорення руху макромолекул, що створює опір у в'язкому середовищі із силою тертя Fо =fV, де V- швидкість руху макромолекул; f- коефіцієнт внутрішнього тертя. Через певний період, коли сили врівноважуються (Fе= Fо), макромолекули рухаються рівномірно з постійною швидкістю V:
Під дією цієї сили відбувається прискорення руху макромолекул, що створює опір у в'язкому середовищі із силою тертя Fо =fV, де V- швидкість руху макромолекул; f- коефіцієнт внутрішнього тертя. Через певний період, коли сили врівноважуються (Fе= Fо), макромолекули рухаються рівномірно з постійною швидкістю V: