
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Биосинтез пуриновых и пиримидиновых нуклеотидов..Содержание книги
Поиск на нашем сайте
пиримидина
пурина
Биосинтез УМФ
КАД-фермент. УМФ и пуриновые нуклеотиды аллостерически ингибируют, а ФРДФ активирует его карбамоилсинтетазную активность, тогда как активность аспартаттранскарбамоилазного домена ингибирует ЦТФ, но активирует АТФ. Этот способ регуляции позволяет предотвратить избыточный синтез не только УМФ, но и всех других пиримидиновых нуклеотидов и обеспечить сбалансированное образование всех четырёх основных пуриновых и пиримидиновых нуклеотидов, необходимых для синтеза РНК.
Репликация Процесс удвоения хромосом называют репликацией. При репликации каждая цепь родительской двухцепочеч-ной ДНК служит матрицей для синтеза новой комплементарной цепи. Вновь образованная двойная спираль имеет одну исходную (родительскую) и одну вновь синтезированную (дочернюю) цепь. Репликацию можно разделить на 4 этапа: образование репликативной вилки (инициация), синтез новых цепей (элонгация), исключение праймеров, завершение синтеза двух дочерних цепей ДНК (терминация). С понятием сайта инициации репликации тесно связано понятие репликон. Репликон — это участок ДНК, который содержит сайт инициации репликации и реплицируется после начала синтеза ДНК с этого сайта. Молекулярные механизмы, которые контролируют количество актов инициации репликации в каждом сайте за один цикл деления клетки, называются контролем копийности. Репликация начинается в сайте инициации репликации с расплетания двойной спирали ДНК, при этом формируется репликационная вилка — место непосредственной репликации ДНК. В каждом сайте может формироваться одна или две репликационные вилки в зависимости от того, является ли репликация одно- или двунаправленной. Более распространена двунаправленная репликация. В каждой репликативной вилке идёт одновременно синтез двух новых (дочерних) цепей. Направление синтеза цепи ДНК совпадает с направлением движения репликативной вилки лишь для одной из вновь синтезируемых цепей (лидирующая цепь). На второй матричной цепи синтез дочерней ДНК осуществляется двумя ферментами: ДНК-полимеразой α и ДНК-полимеразой ε в направлении 5'→3', но против движения репликативной вилки. Поэтому вторая цепь синтезируется прерывисто, короткими фрагментами, которые называют "фрагменты Оказаки" (по имени открывшего их исследователя). Дочерняя цепь ДНК, синтез которой происходит фрагментами, называют отстающей цепью. Каждый фрагмент Оказаки, примерно 100 нуклеотидных остатков, содержит праймер. Праймеры удаляет ДНК-полимераза β, постепенно отщепляя с 3'-конца фрагмента по одному ри-бонуклеотиду. К ОН-группе на 3'-конце предыдущего фрагмента ДНК-полимераза β присоединяет дезоксирибонуклеотиды в количестве, равном вырезанному праймеру и таким образом заполняет брешь, возникающую при удалении рибонуклеотидов.
Фермент ДНК-лигаза катализирует образование фосфодиэфирной связи между 3'-ОН-группой дезоксирибозы одного фрагмента цепи ДНК и 5'-фосфатом следующего фрагмента. Реакция протекает с затратой энергии АТФ. Таким образом, из множества фрагментов Оказаки образуется непрерывная цепь ДНК. В репликационной вилке ДНК копирует крупный белковый комплекс (реплисома), ключевым ферментом которого является ДНК-полимераза. Репликационная вилка движется со скоростью порядка 100 000 пар нуклеотидов в минуту у прокариот и 500—5000 — у эукариот Терминация. необходимо удалить праймер это осуществляется фермент экзонуклеаза, которая садится на гибридные участки и разрушает праймер. Образуются бреши, которые нужно достроить это осуществляется либо пол-α, либо пол-β застройка идет от 3’ к 5’ концу. Возникает проблема сшивания сахарафосфатного остова, эта проблема решается при помощи фермента лигазы, которая может образовывать ковалентные связи с затравкой с помощью энергии АТФ. На ДНК одновременно может существовать несколько репликаторов, в которых начинается инициация репликации, тот участок ДНК, на котором заканчивается синтез, называется терминатор. Участок молекулы ДНК между двумя терминаторами, которые содержат автономную репликационную последовательность, называются репликоны. Т.о. одна молекула ДНК может содержать много репликонов. На концевых участках ДНК возникает проблема с застройкой бреши на лидирующей цепи. С каждой репликацией молекула ДНК укорачивается, концевые участки ДНК не содержат гены и, следовательно, не несут генетической информации. Количество клеточных делений ограничено. В организме есть клетки, которые могут делится неограниченное количество раз. Это предшественники половых клеток, стволовые клетки крови, раковые клетки. Оказалось, что в тканях таких клеток в активной форме находятся фермент теломераза. Теломераза способна удлинять 3’ конец матрицы.
Транскрипия ТРАНСКРИПЦИЯ биосинтез РНК на матрице ДНК; Это матричный процесс в ходе, которого на матрице одноцепочечной ДНК синтезируется одноцепочечная РНК (для всех клеточных организмов), у вирусов возможны варианты: на матриц РНК синтезируется копия РНК. Существуют единицы транскрипции, которые были названы транскриптонами и которые гораздо короче репликонов. Каждый транскриптон включает в себя 3 участка:Инициатор,Кодирующая область,Терминатор.
Ферментом, который обеспечивает процесс транскрипции, является ДНК-зависимая-РНК-полимераза. У проккориот существует один универсальный фермент РНК-полимераза, которая работает на всех транскриптонах. У эукориот существует три варианта полимераз: полимераза 1, полимераза 2, полимераза 3, которые осуществляют различные процессы: полимераза 1 работает на транскрибтонах, которые отвечают за синтез большой рибосомальной РНК; полимераза 2 работает на транскрибтонах, которые отвечают за синтез матричной РНК; полимераза 3 работает на транскрибтонах, которые отвечают за синтез тРНК и малой рибосомальной РНК. Для большой рибосомальной РНК характерен коэффициент 45S, для малой рибосомальной РНК 5S. В настоящее время считается, что у эукориот сначала была одна полимераза 2, а затем из нее за счет модификаций возникли полимераза 1 и полимераза 3. для нормального осуществления транскрипции полимераза должна связываться с определенными белками, которые получили название транскрибирующие факторы. Существует у эукориот по крайне мере несколько транскрибирующих факторов: ТФ1; ТФ2; ТФ3… процесс энергозависим, а как все матричные процессы делится на инициацию, элонгацию и терминацию. 1. инициация начинается с района, который получил название инициатора. Он делится на два отдела:регулятор, промотор. Регулятор необходим для присоединения регуляторных молекул, которые могут активировать или тормозить процесс транскрипции. К промотору в начале транскрипции присоединяется транскрибирующий фактор и только после этого комплекс промотор + Тф опознается РНК-полимераза. Для промотора характерно наличие так называемого ТАТА-бокса. После образования комплекса промотор+ТФ к нему присоединяется РНК-полимераза. Она обладает рядом особенностей: В качестве субстрата используются рибонуклеазы; Может сама начинать синтез, построение «затравки» не нужно; Способна наращивать только 3’ конец, т.е. работать в направлении от 5’ к 3’; Обладает хеликазной активностью.
Присоединение ТФ необходимо для того, чтобы РНК-полимераза могла: опознать промотор, сойти с промотора на кодирующую область. После того как РНК-полимераза сходит на кодирующую область процесс инициации заканчивается. ТФ может остаться в ТАТА-боксе, либо сходит с ТАТА-бокса.
2. элонгация заключается в построении молекулы РНК по правилу комплиментарности от 5’ к 3’ концу. Между дезоксирибонуклеотидами и рибонуклеатидами нет полного соответствия, они слабо реагируют друг с другом и не образуют прочных водородных связей, поэтому копия практически сразу же сходит с матрицы. По мере прохождения РНК-полимераза ДНК восстанавливает свою нормальную структуру в виде двойной спирали.
3. терминация. Терминатор содержит специальную последовательность нуклеотидов, которые могут связываться с терминирующим фактором. У эукориот таких факторов много, у прокариот ρ-фактор, ξ-фактор. Присоединение терминирующих факторов приводит к образованию шпилек на ДНК и РНК-полимераза не может двигаться дальше. Она сходит с первичного транскриптона, и затем они диссоциирует. Для синтеза молекулы РНК используется только одна цепь ДНК в транскриптоне. Выбор цепи определяется положении промотора. Первичные транскрипты нефункциональны и поэтому подвергаются различным модификациям. В общем виде они получили название процессинг или созревание РНК.
ü Вся совокупность ядерных транскриптов РНК-полимеразы II известна как гетерогенная ядерная РНК (гяРНК), поскольку одна из основных характеристик, отличающих эту фракцию ядерных РНК - это чрезвычайно высокая вариабельность размеров входящих в нее транскриптов. По мере синтеза эти транскрипты ковалентно модифицируются по 5'-концам и 3'-концам таким образом, что они становятся отличными от транскриптов, синтезированных другими РНК-полимеразами. Эти модификации будут позже использованы в цитоплазме как сигналы того, что данные информационные РНК должны быть транслированы в белки.
|
|||||||
|
|
Последнее изменение этой страницы: 2016-04-19; просмотров: 817; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.148.115.40 (0.009 с.) |




 синтез ЦТФ из УТФ
синтез ЦТФ из УТФ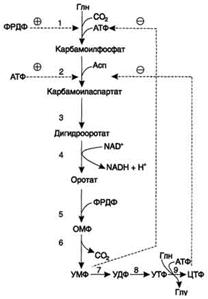 Регуляция синтеза пиримидиновых нуклеотидов. КАД-фермент катализирует реакции 1, 2, 3; дигидрооротатдегидрогеназа - реакцию 4; УМФ синтетаза - реакции 5 и 6; НМФ киназа - реакцию 7; НДФ киназа - реакцию 8; ЦТФ синтетаза - реацию 9.
Регуляция синтеза пиримидиновых нуклеотидов. КАД-фермент катализирует реакции 1, 2, 3; дигидрооротатдегидрогеназа - реакцию 4; УМФ синтетаза - реакции 5 и 6; НМФ киназа - реакцию 7; НДФ киназа - реакцию 8; ЦТФ синтетаза - реацию 9.


