Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Генетическая инженерия дрожжейСодержание книги
Поиск на нашем сайте
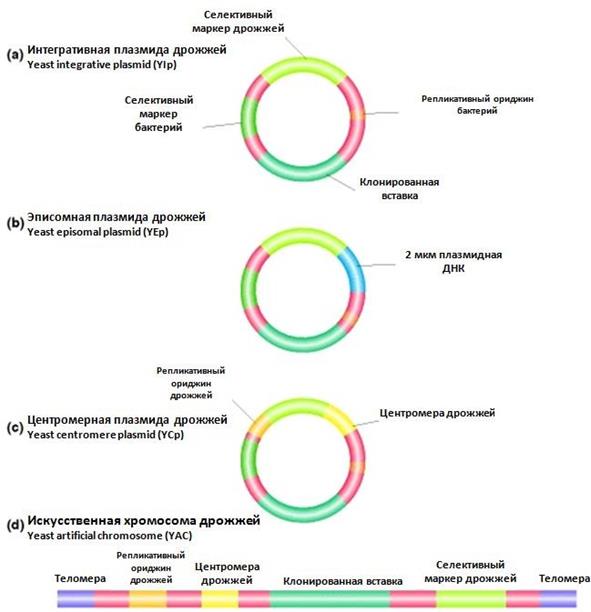
Дрожжевые клетки для экспрессии эукариотических генов используют по следующим причинам. Во-первых, дрожжи – это одноклеточные эукариоты, генетика и физиология которых хорошо изучены, геном дрожжей секвенирован, их можно культивировать в условиях лаборатории и в биореакторах. Во-вторых, в качестве векторных молекул могут использоваться природные плазмиды дрожжей, также клонированы и охарактеризованы сильные дрожжевые промоторы. В-третьих, в клетках дрожжей пост- трансляционные модификации аминокислотных остатков белков (гликозилирование, формирование -S-S- мостиков и др.) осуществляются как у эукариот, прокариоты из-за отсутствия соответствующих ферментных систем не способны проводить такие реакции. Четвертое, дрожжи способны секретировать гетерологичные белки в среду, что облегчает процесс очистки. В-пятых, дрожжи используются в пищевой промышленности и признаны безопасными микроорганизмами. Поэтому процедура допуска при использовании дрожжей для получения медицинских препаратов, существенно упрощена. Также важно, что компоненты и продукты метаболизма дрожжей признаны нетоксичными для человека. Для работы с дрожжами используют различные вектора клонирования. Большинство из них – бинарные векторы, которые реплицируются в дрожжах и бактериях. При эт омс основные манипуляции удобнее проводить в клетках, например, E.coli, а конечные конструкции переносить в дрожжи, чтобы использовать свойства этого хозяина, например, гликозилирование. В основе большинства дрожжевых векторов - природные дрожжевые плазмиды. Они называются 2-мкм (2µ)-плазмиды, они имеют одну дрожжевую ori -последовательность, их содержат большинство штаммов дрожжей. Размер этих плазмид составляет в среднем 6.3 кб, количество достигает 50 копий на клетку. Типичный вектор на основе дрожжевой плазмиды – это вектор экспрессии. Дрожжевые клетки не способны эффективно вырезать интроны, поэтому для клонирования в дрожжевые плазмиды используют либо кДНК, либо ДНК, синтезируемую химическим путем. кДНК встраивают между дрожжевым промотором и дрожжевым сигналом терминации транскрипции (терминатором). В качестве маркерного гена дрожжевой вектор экспрессии содержит собственный ген биосинтеза лейцина (LЕU2). Этим вектором трансформируют штаммы дрожжей, не способные синтезировать лейцин (штаммы ауксотрофы по лейцину) и высевают на среду без лейцина. Вырастают клетки, содержащие плазмиды. Первый шаг на пути оптимизации экспрессии в дрожжах – это выбор промотора. Дрожжевые промоторы отличаются от бактериальных: у дрожжей ТАТА-бокс расположен значительно дальше (40-120 п. о.) от точки инициации транскрипции синтеза белка (у E. сoli - это область -10). Кроме того, дрожжевые промоторы функционируют вместе с активаторами 100 – 1000 п.о. от сайта инициации транскрипции (+1). Поэтому дрожжевые векторы содержат длинную регуляторную область, до 1 кб. В качестве сильных дрожжевых конститутивных промоторов используют промоторы дрожжевых генов алкогольдедидрогеназы и триозофосфатдегидрогеназы, в качестве сильных индуцибельных промоторов – промоторы генов катаболизма галактозы. Эти промоторы активируются галактозой и репрессируются глюкозой. У высших эукариот сайты терминации транскрипции и полиаденилирования расположены на расстоянии нескольких сотен нуклеотидов друг от друга. У дрожжей иначе: полиаденилирование происходит очень близко к 3’-концу транскрипта. Поэтому для эффективной экспрессии в дрожжах вслед за чужеродным геном встраивают дрожжевой сайт терминации транскрипции. Если за кодирующей областью нет терминирующей последовательности, то не образуется стабильная мРНК из-за отсутствия сайта, подходящего для полиаденилирования. Таким образом, подстыковка дрожжевого терминатора транскрипции необходима для повышения стабильности мРНК. Следующее требование для экспрессии – это эффективная трансляция. Для этого необходимо распознавание факторами инициации участка инициации трансляции AUG-кодона. В бактериях это связывание последовательности Шайно-Дальгарно с комплементарным участком 16SРНК. У дрожжей, как и остальных эукариот, нет таких последовательностей. AUG у дрожжей находится в окружении A и U, тогда как гуанин не встречается на протяжении 20-40 пар оснований, предшествующих кодону. Присутствие гуанина в этой области влечет остановку инициации трансляции. Трансляция также ингибируеться, если в 5’-нетранслируемой области имеются петлевые структуры. Используют следующие основные вектора клонирования. Интегративные плазмиды дрожжей – это бактериальные плазмиды, интегрированные в дрожжевой геном (рис. 19а). Эти вектора не содержат дрожжевую последовательность инициации репликации, в связи с чем они самостоятельно не способны реплицировать в дрожжах и размножаются только в интегрированном в геном дрожжей состоянии. Они содержат небольшие участки, гомологичные фрагментам хромосомной ДНК дрожжей. Интеграция происходит за счет гомологичной рекомбинации с частотой 10-5. Эти плазмиды содержат также маркерные гены (например, ген биосинтеза лейцина LEU), которые обеспечивают селективный отбор в дрожжевых клетках. Основной недостаток в использовании интегративных векторов – это низкий уровень экспрессии (одна копия), поскольку плазмида наследуется как часть дрожжевого генома. Эту проблему решают встраиванием векторных последовательностей в область тандемных генов (гены рРНК), такая интеграция позволяет получить множество копий клонированных участков. Встраивание в такой вектор дрожжевой последовательности инициации репликации из дрожжевой хромосомы, называемой ARS (a utonomously r eplicating s equence), позволило получить дрожжевые репликативные плазмиды. Такая плазмида может реплицироваться самостоятельно в клетках дрожжей без интеграции в хромосому. Проблема в том, что при делении почкованием плазмиды неравномерно распределяются по дочерним клеткам, часто потомки полностью лишены плазмид. Поэтому они не соответствуют для репродуктивной экспрессии.
Рис. 20. Векторные системы дрожжей [Modern Genetic Analysis.Griffiths AJF, Gelbart WM, Miller JH, et al.New York: W. H. Freeman; 1999. www.ncbi.nlm.nih.gov/books/NBK21428/]
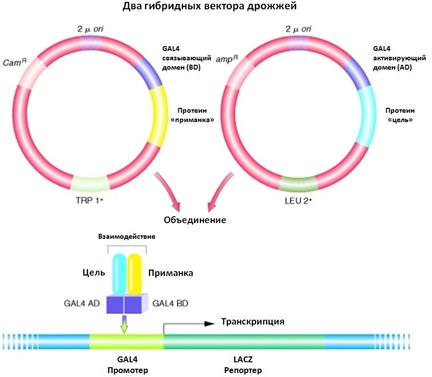
Эписомные плазмиды дрожжей (рис. 19b). Эписома – это генетический элемент, который может существовать в свободном и интегрированном в хромосому состоянии, т.е. как плазмида, а также в виде части хромосомы. Именно такими свойствами обладают эписомные плазмиды. Они созданы на основе дрожжевых 2µ-плазмид. Для получения дрожжевых эписомных плазмид объединяют участок инициации репликации дрожжевой 2µ-плазмиды и интегративные плазмиды дрожжей. Полученные в результате объединения эписомные плазмиды являются многокопийными и позволяют получить от 30 до 50 копий на клетку. Эти плазмиды также неравномерно передаются дочерним клеткам, но стабильно поддерживаются за счет высокой копийности. Эписомные плазмиды применяются для получения высокого уровня экспрессии гетерологичных генов и часто именно они используются для получения генно- инженерных продуктов дрожжей. Центромерные плазмиды дрожжей получают из интегративных или эписомных путем вставки в них последовательности дрожжевой центромеры (CEN-последовательности), (рис. 19c). Центромерная последовательность позволяет плазмидам правильно расходиться при митотическом делении, подобно хромосомам. Эти плазмиды очень стабильны, так как успешно распределяются между дочерними клетками. Их нгедостаток в хромосомноподобном поведении. Что обуславливает низкую копийность (1-3 копии на клетку). Экспрессию такой плазмиды можно повысить за счет оптимизации транскрипции и трансляции. Искусственные дрожжевые хромосомы, YAC (Yeast artificial chromosome), предназначены для клонирования больших фрагментов генома (300 kb), (рис. 19d). Такой вектор содержит сайт инициации репликации дрожжевой хромосомы - ARS, сегмент центромерной области дрожжевой хромосомы – CEN, а также теломерные последовательности дрожжевой хромосомы – TEL- последовательности из повторяющихся олигонуклеотидов на концах линейных молекул ДНК, обеспечивающих стабильность вектора. YAC-системы стабильны, они были использованы при физическом картировании генома человека, при анализе больших транскриптов, при создании геномных библиотек. Важно, что встройка крупных фрагментов позволяет клонировать весь геном в виде относительно небольшого числа гибридных мини-хромосом, которые легко хранить и использовать в последующей работе. При встраивании чужеродной ДНК в YAC-вектор может происходить нарушение рамки считывания маркерного гена, что используется при отборе. Кроме того, некоторые YAC-векторы несут селективный маркер, независимый от сайта клонирования. Двухгибридные системы дрожжей служат для идентификации белок- белковых взаимодействий (рис. 20). Это новая система дрожжей для выявления взаимодействия белков in vivo. Она основана на организации и функционировании регуляторного белка дрожжей GAL4, который является активатором транскрипции генов, кодирующих ферменты метаболизма галактозы. GAL4 состоит из двух доменов: N-концевого домена, который связывается со специфическими последовательностями ДНК, а также С- концевого домена, необходимого для активации транскрипции. Домены белка GAL4 активны при взаимодействии друг с другом.
Рис. 20. Двухгибридная система дрожжей для идентификации генных взаимодействий. Тестирование использует связывание двух белков для восстановления функции GAL4-белка, который активирует репортерный ген. http://www.ncbi.nlm.nih.gov/books/NBK21248/
Когда домены входят в состав двух разных гибридных генов, они способны функционально активироваться in vivo, если сшитые с ними белки взаимодействуют друг с другом. Метод основан на использовании дву- плазмидной системы в дрожжах, основа тестирования – активация биосинтеза галактозидазы. Ген одного из тестируемых белков (Bait-белок) сшивают с ДНК- фрагментом, соответствующим ДНК-связывающему домену GAL4 (BD-домен) на одной плазмиде, ген другого (Target-белок) – с фрагментом ДНК, кодирующим активаторный домен GAL4 (AD-домен), на другой плазмиде. Обе плазмиды котрансформируют в клетки дрожжей. Если в результате экспрессии фьюжен-конструкций тестируемые белки (Bait и Target) взаимодействуют, это приводит к физическому взаимодействию AD и BD доменов и активации синтеза галактозидазы, активации гена LACZ. При отсутствии взаимодействия, экспрессии GAL4 не происходит. Дрожжевая двухгибридная система нашла широкое применение для изучения взаимодействия белков различных организмов и вирусов. Рекомбинантные белки, синтезированные в S. сerevisiae, применяются в качестве вакцин, фармацевтических препаратов и для диагностики. Так, для формирования иммунитета используют генно-инженерную вакцину против вируса гепатита В, созданную на основе антигена вируса – белка оболочки. Для диагностики используют белок вируса гепатита С, а также антигены HIV-1.В качестве лекарственых средств используют инсулин, фактор роста, фактор XIIIa системы свертывания крови и др. При использовании эписомного вектора дрожжей с дрожжевой сигнальной последовательностью получен белок гирудин, кодируемый медицинской пиявкой Hirudo medicinals. Белок – мощный антикоагулянт крови и не вызывает иммунологичнеских реакций у человека, способен разрушать сгустки венозной крови и устранять проявления тромбоза. На эписомном векторе клониролван ген человека супероксид-дисмутаза, который катализирует связывание супероксид-аниона с образованием перекиси водорода, предотвращая повреждение клеток. Препарат активно используется при хирургических вмешательствах, лечении остеоартрита, ревматоидного артрита, склеродермии и др. Однако, при работе с рекомбинантными дрожжами обнаружились проблемы. Прежде всего уровень экспрессии дрожжевых систем низкий и уступает бактериальным системам экспрессии. При масштабировании плазмиды теряются, а рекомбинантные белки часто гипергликозилированы, что влияет на иммуногенность. Кроме того, в некоторых системах секретируемые белки концентрируются в периплазме, что усложняет очистку. В практике используются и другие дрожжевые системы экспрессии. Например, метилотрофные дрожжи Pichia pastoris в качестве организма-хозяина, которые без больших затрат можно выращивать в промышленных биореакторах. С использованием интегративного вектора в этих дрожжах получают поверхностный антиген вируса гепатита В. На их основе был осуществлен синтез бычьего лизоцима С2. Это желудочный фермент, разрушающий клеточные стенки бактерий, он устойчив к пртеазам и сохраняет активность в узком диапазоне рН, что позволяет использовать его в качестве добавки к кормам жвачных животных для улучшения пищеварения.В результате при ферментации 10 л культуры P. Pastoris в течении 200 ч в непрерывном режиме при высокой плотности клеток синтезировалось примерно 20 г лизоцима С2. Аутентичные гетерологичные белки были получены и с помощью других дрожжевых систем. Например, гемоглобин А человека с правильной тетрамерной структурой альфа- и бета-цепей, а также человеческая урокиназа получены в дрожжах H. polymorpha и S. сerevisiae. Таким образом, дрожжевые системы экспрессии играют важную роль в получении гетерологичных белков для промышленных и медицинских целей.
|
||||
|
|
Последнее изменение этой страницы: 2022-09-03; просмотров: 370; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.48.72 (0.01 с.) |