Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Гипотезы о происхождении вирусовСтр 1 из 5Следующая ⇒
Содержание · 1Этимология названия · 2История исследований · 3Происхождение o 3.1Гипотезы о происхождении вирусов § 3.1.1Регрессивная гипотеза § 3.1.2Гипотеза клеточного происхождения § 3.1.3Гипотеза коэволюции o 3.2Мир РНК o 3.3Прионы · 4Биология o 4.1Вирусы как форма жизни o 4.2Структура § 4.2.1Капсид § 4.2.1.1Спиральный § 4.2.1.2Икосаэдрический § 4.2.1.3Продолговатый § 4.2.1.4Комплексный § 4.2.2Оболочка o 4.3Геном o 4.4Жизненный цикл § 4.4.1Особенности жизненного цикла различных групп o 4.5Действие на клетки o 4.6Диапазон хозяев § 4.6.1Распространение o 4.7Эволюция · 5Классификация o 5.1Классификация ICTV o 5.2Классификация по Балтимору · 6Роль в заболеваниях человека o 6.1Эпидемиология o 6.2Эпидемии и пандемии o 6.3Злокачественные опухоли o 6.4Защитная реакция хозяина o 6.5Профилактика и лечение § 6.5.1Вакцины § 6.5.2Противовирусные препараты · 7Вирусные заболевания у различных организмов o 7.1Вирусы животных § 7.1.1Вирусы беспозвоночных o 7.2Вирусы растений o 7.3Вирусы грибов o 7.4Вирусы протистов o 7.5Вирусы бактерий o 7.6Вирусы архей o 7.7Вирусы вирусов · 8Роль вирусов в биосфере o 8.1Роль в водных экосистемах · 9Роль в эволюции · 10Применение o 10.1В науках о жизни и медицине o 10.2В материаловедении и нанотехнологиях o 10.3Искусственные вирусы o 10.4Вирусы как оружие · 11В массовой культуре o 11.1В литературе o 11.2В кинематографе o 11.3В мультипликации · 12См. также · 13Примечания · 14Литература · 15Ссылки Этимология названия Слово «вирус» образовано от лат. virus — «яд»[2]. Для обозначения агента, способного вызывать инфекционную болезнь, оно впервые было применено в 1728 году[11], ещё до открытия вирусов, сделанного в 1892 году Дмитрием Ивановским; им же был введён термин фильтрующийся вирус как обозначение небактериального болезнетворного агента, способного проходить сквозь бактериальные фильтры — фильтроваться. Термин «вирион», создание которого датируется 1959 годом[12], применяется для обозначения единичной стабильной вирусной частицы, покинувшей клетку и способной инфицировать другие клетки того же типа[13]. История исследований
Мартин Бейеринк в своей лаборатории в 1921 году По мере накопления данных об инфекционных заболеваниях различных организмов стало очевидно, что далеко не все из них вызываются патогенами, известными на тот момент — бактериями, протистами или микроскопическими грибками. В частности, Луи Пастер не смог найти агент, вызывающий бешенство, и предполагал, что этот патоген слишком мал, чтобы увидеть его в микроскоп[14]. В 1884 году французский микробиолог Шарль Шамберлан изобрёл фильтр (сейчас известный как фильтр Шамберлана или фильтр Шамберлана—Пастера), поры которого меньше бактерий. С помощью этого фильтра можно полностью удалить бактерии из раствора[15]. В 1892 году русский биолог Дмитрий Ивановский использовал его для изучения вида, сейчас известного как вирус табачной мозаики. Его эксперименты показали, что экстракт перетёртых листьев заражённых растений табака сохраняет инфекционные свойства и после фильтрации. Ивановский предположил, что инфекция может вызываться токсином, выделяемым бактериями, однако он не развил эту идею[16]. В то время считалось, что любой инфекционный агент можно выделить на фильтре и выращивать в питательной среде — таков один из постулатов микробной теории болезней[6]. Кроме того, Ивановский в оптическом микроскопе наблюдал в заражённых клетках растений кристаллоподобные тела, которые в современном понимании являлись скоплениями вирусов, впоследствии они были названы «кристаллами Ивановского»[17]. В 1898 году голландский микробиолог Мартин Бейеринк повторил эксперименты Ивановского и пришёл к выводу, что прошедший сквозь фильтр инфекционный материал есть не что иное, как новая форма инфекционных агентов[18]. Он подметил, что агент размножался только в делящихся клетках, однако его опыты не выявили того, что он представляет собой частицы. Бейеринк назвал его Contagium vivum fluidum (дословно лат. растворимый живой микроб) и вновь ввёл в употребление слово «вирус»[16]. Он утверждал, что по своей природе вирус жидкий. Эта теория впоследствии была опровергнута Уэнделлом Стэнли, доказавшим, что вирусы представляют собой частицы[16]. В том же году Фридрих Лёффлер и Пауль Фрош обнаружили первый вирус животных — возбудитель ящура (афтовирус), пропустив его через схожий фильтр[19].
В начале XX века английский бактериолог Фредерик Туорт открыл группу вирусов, инфицирующих бактерии (сейчас они известны как бактериофаги[20] или просто фаги), а франко-канадский микробиолог Феликс Д’Эрелль описал вирусы, которые при добавлении к бактериям на агаре образуют вокруг себя пространство с мёртвыми бактериями. Д’Эрелль сделал точные разведения суспензии этих вирусов и установил наибольшее разведение (наименьшую концентрацию вирусов), при котором не все бактерии погибают, но тем не менее образуются отдельные области с мёртвыми клетками. Подсчитав число таких областей и учитывая коэффициент разведения, он определил число вирусных частиц в первоначальной суспензии[21]. Фаги были объявлены потенциальным средством от болезней, таких как тиф и холера, однако это было забыто в связи с открытием уникальных свойств пенициллина. Изучение фагов дало информацию о явлении «включения» и «выключения» генов, а также позволило использовать их для введения в бактериальный геном чужеродных генов.
К концу XIX века было известно, что вирусы обладают инфекционными свойствами, способны проходить через фильтры и нуждаются в живом хозяине для размножения. В то время вирусы в исследовательских целях культивировали только в растениях и животных. В 1906 году Росс Грэнвилл Гаррисон изобрёл метод выращивания тканей в лимфе, и в 1913 году Штейнард, Израэли и Ламберт использовали этот метод при выращивании вируса осповакцины на фрагментах ткани роговицы морских свинок[22]. В 1928 году Г. Б. Мэйтланд и М. К. Мэйтланд вырастили вирус осповакцины на суспензии из измельчённых куриных почек. Этот метод не применялся широко до конца 1950-х годов, когда в больших масштабах стали выращивать полиовирус для производства вакцины[23]. Другое крупное достижение принадлежит американскому патологу Эрнесту Уильяму Гудпасчеру; в 1939 г он вырастил вирус гриппа и несколько других вирусов в оплодотворённых куриных яйцах[24]. В 1949 году Джон Франклин Эндерс, Томас Уэллер и Фредерик Роббинс вырастили полиовирус на культуре клеток зародыша человека. Это был первый вирус, выращенный не на тканях животных или яйцах. Эта работа дала возможность Джонасу Солку создать эффективную полиовакцину (вакцину против полиомиелита)[25]. Первые изображения вирусов были получены после изобретения электронного микроскопа немецкими инженерами Эрнстом Руской и Максом Кноллем[26]. В 1935 году американский биохимик и вирусолог Уэнделл Мередит Стэнли тщательно изучил вирус табачной мозаики и обнаружил, что он по большей части состоит из белка[27]. Спустя короткое время этот вирус был разделён на белковую и РНК-составляющую[28]. Вирус табачной мозаики был кристаллизован первым среди вирусов, что позволило многое узнать о его структуре. Первая рентгенограмма кристаллизованного вируса была получена Берналем и Фэнкухеном в конце 1930-х годов. На основании полученных ею изображений Розалинд Франклин в 1955 году определила полную структуру вируса[29]. В том же году Хайнц Френкель-Конрат и Робли Уилльямс показали, что очищенная РНК вируса табачной мозаики и белок оболочки способны к самосборке в функциональный вирус. Это позволило им предположить, что подобный механизм лежит в основе сборки вирусов внутри клеток-хозяев[30]. Вторая половина XX века стала периодом расцвета вирусологии. В то время было открыто свыше 2000 видов вирусов животных, растений и бактерий[31]. В 1957 году были открыты лошадиный артеривирус и возбудитель вирусной диареи коров (пестивирус). В 1963 году Барух Бламберг открыл вирус гепатита B[32], а в 1965 году Хоуард Темин описал первый ретровирус. В 1970 году Темин и Дейвид Балтимор независимо друг от друга описали обратную транскриптазу, ключевой фермент, с помощью которого ретровирусы синтезируют ДНК-копии своих РНК[33]. В 1983 году группа учёных во главе с Люком Монтанье из Института Пастера во Франции впервые выделила ретровирус, известный сейчас как ВИЧ[34].
В 2002 году в Нью-Йоркском университете был создан первый синтетический вирус (вирус полиомиелита)[35]. Происхождение Появление вирусов на эволюционном древе жизни неясно: некоторые из них могли образоваться из плазмид, небольших молекул ДНК, способных передаваться от одной клетки к другой, в то время как другие могли произойти от бактерий. В эволюции вирусы являются важным звеном горизонтального переноса генов, обуславливающего генетическое разнообразие[36]. Некоторые учёные считают вирусы особой формой жизни, так как они имеют генетический материал, способны создавать себе подобные вирусы, и эволюционируют путем естественного отбора. Однако у вирусов отсутствуют важные характеристики (такие как клеточное строение и собственный обмен веществ), без которых их нельзя отнести к живому. Так как они обладают некоторыми, но не всеми свойствами живого, вирусы описываются как «организмы на краю жизни». Вирусы найдены везде, где есть жизнь, и, вероятно, вирусы существуют с момента появления первых живых клеток[37]. Происхождение вирусов неясно, поскольку они не оставляют каких бы то ни было ископаемых остатков, а их родственные связи можно изучать только методами молекулярной филогенетики[38]. Регрессивная гипотеза Согласно этой гипотезе, вирусы когда-то были мелкими клетками, паразитирующими в более крупных клетках. С течением времени эти клетки предположительно утратили гены, которые были «лишними» при паразитическом образе жизни. Эта гипотеза основывается на наблюдении, что некоторые бактерии, а именно риккетсии и хламидии, представляют собой клеточные организмы, которые, тем не менее, подобно вирусам могут размножаться только внутри другой клетки. Эту гипотезу также называют гипотезой дегенерации [41][42] или гипотезой редукции [43].
Гипотеза коэволюции Эта гипотеза предполагает, что вирусы возникли из сложных комплексов белков и нуклеиновых кислот в то же время, что и первые на Земле живые клетки, и зависят от клеточной жизни вот уже миллиарды лет. Помимо вирусов, существуют и другие неклеточные формы жизни. Например, вироиды — это молекулы РНК, которые не рассматриваются как вирусы, потому что у них нет белковой оболочки. Тем не менее, ряд характеристик сближает их с некоторыми вирусами, а потому их относят к субвирусным частицам[47]. Вироиды являются важными патогенами растений[48]. Они не кодируют собственные белки, однако взаимодействуют с клеткой-хозяином и используют её для осуществления репликации своей РНК[49]. Вирус гепатита D имеет РНК-геном, схожий с геномом вироидов, однако сам не способен синтезировать белок оболочки. Для формирования вирусных частиц он использует белок капсида вируса гепатита B и может размножаться только в клетках, заражённых этим вирусом. Таким образом, вирус гепатита D является дефектным вирусом[50]. Вирофаг Спутник схожим образом зависит от мимивируса, поражающего простейшее Acanthamoeba castellanii [51]. Эти вирусы зависят от присутствия в клетке-хозяине другого вируса и называются вирусами-сателлитами. Подобные вирусы демонстрируют, как может выглядеть промежуточное звено между вирусами и вироидами[52][53]. Каждая из этих гипотез имеет свои слабые места: регрессивная гипотеза не объясняет, почему даже мельчайшие клеточные паразиты никак не походят на вирусы. Гипотеза побега не даёт объяснения появлению капсида и других компонентов вирусной частицы. Гипотеза коэволюции противоречит определению вирусов как неклеточных частиц, зависимых от клеток-хозяев[43]. Тем не менее, в настоящее время многие специалисты признают вирусы древними организмами, появившимися, предположительно, ещё до разделения клеточной жизни на три домена[54]. Это подтверждается тем, что некоторые вирусные белки не обнаруживают гомологии с белками бактерий, архей и эукариот, что свидетельствует о сравнительно давнем обособлении этой группы. В остальном же достоверно объяснить происхождение вирусов на основании трёх закрепившихся классических гипотез не удаётся, что делает необходимыми пересмотр и доработку этих гипотез[54]. Мир РНК Гипотеза мира РНК[55] и компьютерный анализ последовательностей вирусной ДНК и ДНК хозяина дают лучшее понимание эволюционных взаимоотношений между различными группами вирусов и могут помочь определить предков современных вирусов. До настоящего времени такие исследования пока не прояснили, какая из трёх основных гипотез верна[55]. Однако представляется маловероятным, чтобы все современные вирусы имели общего предка, и, возможно, в прошлом вирусы независимо возникали несколько раз по одному или нескольким механизмам, так как между различными группами вирусов имеются значительные различия в организации генетического материала[56].
Препарат головного мозга коровы, поражённой коровьим бешенством. В сером веществе образуются микроскопические полости, которые придают ткани вид губки Прионы Основная статья: Прионы Прионы — это инфекционные белковые молекулы, не содержащие ДНК или РНК[57]. Они вызывают такие заболевания, как почесуха овец[58], губчатая энцефалопатия крупного рогатого скота и хроническая слабость (англ. chronic wasting disease) у оленей. К прионным болезням человека относят куру, болезнь Крейтцфельдта — Якоба и синдром Герстмана — Штраусслера — Шейнкера[59]. Прионы способны стимулировать образование собственных копий. Прионный белок способен существовать в двух изоформах: нормальной (PrPC) и прионной (PrPSc). Прионная форма, взаимодействуя с нормальным белком, способствует его превращению в прионную форму. Хотя прионы фундаментально отличаются от вирусов и вироидов, их открытие даёт больше оснований поверить в то, что вирусы могли произойти от самовоспроизводящихся молекул[60]. Биология Вирусы как форма жизни Пока вирус находится во внеклеточной среде или в процессе заражения клетки, он существует в виде независимой частицы. Вирусные частицы (вирионы) состоят из двух или трёх компонентов: генетического материала в виде ДНК или РНК (некоторые, например мимивирусы, имеют оба типа молекул); белковой оболочки (капсида), защищающей эти молекулы, и, в некоторых случаях, — дополнительных липидных оболочек. Наличие капсида отличает вирусы от вирусоподобных инфекционных нуклеиновых кислот — вироидов. В зависимости от того, каким типом нуклеиновой кислоты представлен генетический материал, выделяют ДНК-содержащие вирусы и РНК-содержащие вирусы; на этом принципе основана классификация вирусов по Балтимору. Ранее к вирусам также ошибочно относили прионы, однако впоследствии оказалось, что эти возбудители представляют собой особые инфекционные белки и не содержат нуклеиновых кислот. Форма вирусов варьирует от простой спиральной и икосаэдрической до более сложных структур. Размеры среднего вируса составляют около одной сотой размеров средней бактерии. Большинство вирусов слишком малы, чтобы быть отчётливо различимыми под световым микроскопом. Вирусы являются облигатными паразитами, так как не способны размножаться вне клетки. Вне клетки вирусные частицы не проявляют признаки живого и ведут себя как частицы биополимеров. От живых паразитарных организмов вирусы отличаются полным отсутствием основного и энергетического обмена и отсутствием сложнейшего элемента живых систем — аппарата трансляции (синтеза белка), степень сложности которого превышает таковую самих вирусов. Согласно одному из определений вирусы представляют собой форму жизни, согласно другому вирусы являются комплексами органических молекул, взаимодействующими с живыми организмами. Вирусы характеризуют как «организмы на границе живого»[19]. Вирусы похожи на живые организмы в том, что они имеют свой набор генов и эволюционируют путём естественного отбора[61], а также в том, что способны размножаться, создавая собственные копии путём самосборки. Вирусы имеют генетический материал, однако лишены клеточного строения, а именно эту черту обычно рассматривают как фундаментальное свойство живой материи. У вирусов нет собственного обмена веществ, и для синтеза собственных молекул им необходима клетка-хозяин. По этой причине они не способны размножаться вне клетки[62]. В то же время такие бактерии, как риккетсии и хламидии, несмотря на то, что не могут размножаться вне клеток хозяина, считаются живыми организмами[63][64]. Общепризнанные формы жизни размножаются делением клетки, в то время как вирусные частицы самопроизвольно собираются в инфицированной клетке. От роста кристаллов размножение вирусов отличается тем, что вирусы наследуют мутации и находятся под давлением естественного отбора. Самосборка вирусных частиц в клетке даёт дополнительное подтверждение гипотезы, что жизнь могла зародиться в виде самособирающихся органических молекул[3]. Опубликованные в 2013 году данные о том, что некоторые бактериофаги обладают собственной иммунной системой, способной к адаптации[65], являются дополнительным доводом в пользу определения вируса как формы жизни. Структура
Вирусы демонстрируют огромное разнообразие форм и размеров. Как правило, вирусы значительно мельче бактерий. Большинство изученных вирусов имеют диаметр в пределах от 20 до 300 нм. Некоторые филовирусы имеют длину до 1400 нм, но их диаметр составляет лишь 80 нм[66]. В 2013 году самым крупным из известных вирусов считался Pandoravirus размерами 1 × 0,5 мкм, однако в 2014 году из многолетней мерзлоты из Сибири был описан Pithovirus, достигающий 1,5 мкм в длину и 0,5 мкм в диаметре. В настоящий момент он считается крупнейшим из известных вирусов[67]. Большинство вирионов невозможно увидеть в световой микроскоп, поэтому используют электронные — как сканирующие, так и просвечивающие[68]. Чтобы вирусы резко выделялись на окружающем фоне, применяют электронноплотные «красители». Они представляют собой растворы солей тяжёлых металлов, таких как вольфрам, которые рассеивают электроны на покрытой ими поверхности. Однако обработка такими веществами ухудшает видимость мелких деталей. В случае негативного контрастирования «окрашивается» только фон[69]. Зрелая вирусная частица, известная как вирион, состоит из нуклеиновой кислоты, покрытой защитной белковой оболочкой — капсидом. Капсид складывается из одинаковых белковых субъединиц, называемых капсомерами [70]. Вирусы могут также иметь липидную оболочку поверх капсида (суперкапсид), образованную из мембраны клетки-хозяина. Капсид состоит из белков, кодируемых вирусным геномом, а его форма лежит в основе классификации вирусов по морфологическому признаку[71][72]. Сложноорганизованные вирусы, кроме того, кодируют специальные белки, помогающие в сборке капсида. Комплексы белков и нуклеиновых кислот известны как нуклеопротеины, а комплекс белков вирусного капсида с вирусной нуклеиновой кислотой называется нуклеокапсидом. Форму капсида и вириона в целом можно механически (физически) исследовать при помощи сканирующего атомно-силового микроскопа[73][74]. Капсид Основная статья: Капсид Классифицируют четыре морфологических типа капсидов вирусов: спиральный, икосаэдрический, продолговатый и комплексный. Спиральный Эти капсиды состоят из одного типа капсомеров, уложенных по спирали вокруг центральной оси. В центре этой структуры может находиться центральная полость или канал. Такая организация капсомеров приводит к формированию палочковидных и нитевидных вирионов: они могут быть короткими и очень плотными или длинными и очень гибкими. Генетический материал, как правило, представлен одноцепочечной РНК (в некоторых случаях одноцепочечной ДНК) и удерживается в белковой спирали ионными взаимодействиями между отрицательными зарядами на нуклеиновых кислотах и положительными зарядами на белках. В целом, длина спирального капсида зависит от длины окружённой им нуклеиновой кислоты, а диаметр определяется размером и расположением капсомеров. Примером спирального вируса может служить вирус табачной мозаики[75].
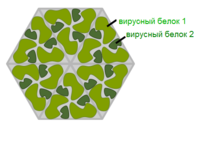
Вирус с капсомером типа гексон. Эта иллюстрация показывает, что вирусный капсид может быть выстроен из множественных копий всего лишь двух белков Икосаэдрический Большинство вирусов животных имеют икосаэдрическую или почти шарообразную форму с икосаэдрической симметрией. Правильный икосаэдр является оптимальной формой для закрытого капсида, сложенного из одинаковых субъединиц. Минимальное необходимое число одинаковых капсомеров — 12, каждый капсомер состоит из пяти идентичных субъединиц. Многие вирусы, такие как ротавирус, имеют более двенадцати капсомеров и выглядят круглыми, но сохраняют икосаэдрическую симметрию. Капсомеры, находящиеся в вершинах, окружены пятью другими капсомерами и называются пентонами. Капсомеры треугольных граней имеют 6 соседей-капсомеров и называются гексонами [76]. Гексоны, по существу, являются плоскими, а пентоны, образующие 12 вершин, — изогнутыми. Один и тот же белок может выступать субъединицей и пентомеров, и гексамеров, или же они могут состоять из различных белков. Продолговатый Продолговатыми называют икосаэдрические капсиды, вытянутые вдоль оси симметрии пятого порядка. Такая форма характерна для головок бактериофагов[77]. Комплексный Форма этих капсидов ни чисто спиральная, ни чисто икосаэдрическая. Они могут нести дополнительные наружные структуры, такие как белковые хвосты или сложные наружные стенки. Некоторые бактериофаги, такие как фаг Т4, имеют комплексный капсид, состоящий из икосаэдрической головки, соединённой со спиральным хвостом, который может иметь шестигранное основание с отходящими от него хвостовыми белковыми нитями. Этот хвост действует наподобие молекулярного шприца, прикрепляясь к клетке-хозяину и затем впрыскивая в неё генетический материал вируса[78].
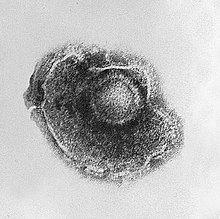
Вирус герпеса, окружённый липидной оболочкой (суперкапсидом) Оболочка Некоторые вирусы окружают себя дополнительной оболочкой из модифицированной клеточной мембраны (плазматической или внутренней, такой как ядерная мембрана или мембрана эндоплазматического ретикулума). Этот дополнительный билипидный слой называется суперкапсидом, а шиповидные выступы на нём — пепломерами. Липидная оболочка вируса испещрена белками, кодируемыми вирусными геномом и геномом хозяина; сама же мембрана, а также любые её углеводные компоненты происходят целиком из клетки-хозяина. Таким образом формируют свою оболочку вирус гриппа и ВИЧ. Инфекционность большинства вирусов, имеющих оболочку, зависит именно от этой оболочки[79]. Поксвирусы представляют собой крупные сложноорганизованные вирусы с необычной морфологией. Генетический материал вируса связан с белками в центральной дисковидной структуре, известной как нуклеоид. Нуклеоид окружён мембраной и двумя боковыми телами неизвестной функции. Вирус имеет наружную оболочку с большим количеством белков на её поверхности. Весь вирион слегка плейоморфен (то есть способен изменять форму и размер в зависимости от условий) и может принимать форму от овальной до блоковидной[80]. Мимивирус является одним из крупнейших описанных вирусов и имеет икосаэдрический капсид диаметром 400—500 нм. Белковые филаменты, отходящие от поверхности вириона, достигают 100 нм в длину[81][82]. В 2011 году исследователи обнаружили ещё более крупный вирус на океаническом дне вблизи побережья Чили. Вирус, которому было дано временное название Megavirus chilensis, можно увидеть даже в обычный оптический микроскоп[83]. Геном Жизненный цикл Основная статья: Жизненный цикл вируса
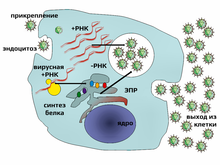
Типичный жизненный цикл вируса на примере вируса гепатита C Вирусы не размножаются клеточным делением, поскольку не имеют клеточного строения. Вместо этого они используют ресурсы клетки-хозяина для образования множественных копий самих себя, и их сборка происходит внутри клетки. Условно жизненный цикл вируса можно разбить на несколько взаимоперекрывающихся этапов (обычно выделяют 6 этапов[96]): · Прикрепление представляет собой образование специфичной связи между белками вирусного капсида и рецепторами на поверхности клетки-хозяина. Это специфичное связывание определяет круг хозяев вируса. Например, ВИЧ поражает только определённый тип человеческих лейкоцитов. Это связано с тем, что оболочечный гликопротеин вируса gp120 специфично связывается с молекулой CD4 — хемокиновым рецептором, который обычно встречается на поверхности CD4+ T-лимфоцитов. Этот механизм обеспечивает инфицирование вирусом только тех клеток, которые способны осуществить его репликацию. Связывание с рецептором может вызвать конформационные изменения белка оболочки (или белка капсида в случае безоболочечного вируса), что в свою очередь служит сигналом к слиянию вирусной и клеточной мембран и проникновению вируса в клетку. · Проникновение в клетку. На следующем этапе вирусу необходимо доставить внутрь клетки свой генетический материал. Некоторые вирусы также переносят внутрь клетки собственные белки, необходимые для её реализации (особенно это характерно для вирусов, содержащих негативные РНК). Различные вирусы для проникновения в клетку используют разные стратегии: например, пикорнавирусы впрыскивают свою РНК через плазматическую мембрану, а вирионы ортомиксовирусов захватываются клеткой в ходе эндоцитоза и попадают в кислую среду лизосом, где происходит депротеинизация вирусной частицы, после чего РНК в комплексе с вирусными белками преодолевает лизосомальную мембрану и попадает в цитоплазму. Вирусы также различают по тому, где происходит их репликация: часть вирусов (например, те же пикорнавирусы) размножается в цитоплазме клетки, а часть (например, ортомиксовирусы) в её ядре. Процесс инфицирования вирусами клеток грибов и растений отличается от инфицирования клеток животных. Растения имеют прочную клеточную стенку, состоящую из целлюлозы, а грибы — из хитина, так что большинство вирусов могут проникнуть в них только после повреждения клеточной стенки[97]. Однако почти все вирусы растений (включая вирус табачной мозаики) могут перемещаться из клетки в клетку в форме одноцепочечных нуклеопротеиновых комплексов через плазмодесмы[98]. Бактерии, как и растения, имеют крепкую клеточную стенку, которую вирусу, чтобы попасть внутрь, приходится повредить. Но в связи с тем, что клеточная стенка бактерий намного тоньше таковой у растений, некоторые вирусы выработали механизм впрыскивания генома в бактериальную клетку через толщу клеточной стенки, при котором капсид остаётся снаружи[99]. · Лишение оболочек представляет собой процесс потери капсида. Это достигается при помощи вирусных ферментов или ферментов клетки-хозяина, а может быть и результатом простой диссоциации. В конечном счёте вирусная геномная нуклеиновая кислота освобождается. · Репликация вирусов подразумевает, прежде всего, репликацию генома. Репликация вируса включает синтез мРНК ранних генов вируса (с исключениями для вирусов, содержащих положительную РНК), синтез вирусных белков, возможно, сборку сложных белков и репликацию вирусного генома, которая запускается после активации ранних или регуляторных генов. Вслед за этим может последовать (у комплексных вирусов с крупными геномами) ещё один или несколько кругов дополнительного синтеза мРНК: «поздняя» экспрессия генов приводит к синтезу структурных или вирионных белков.
Отпочковывание вируса от клетки-хозяина · Вслед за этим происходит сборка вирусных частиц, позже происходят некоторые модификации белков. У вирусов, таких как ВИЧ, такая модификация (иногда называемая созреванием) происходит после выхода вируса из клетки-хозяина[100]. · Выход из клетки. Вирусы могут покинуть клетку после лизиса, процесса, в ходе которого клетка погибает из-за разрыва мембраны и клеточной стенки, если такая есть. Эта особенность есть у многих бактериальных и некоторых животных вирусов. Некоторые вирусы подвергаются лизогенному циклу, где вирусный геном включается путём генетической рекомбинации в специальное место хромосомы клетки-хозяйки. Тогда вирусный геном называется провирусом, или, в случае бактериофага, профагом [101]. Когда клетка делится, вирусный геном также удваивается. В пределах клетки вирус в основном не проявляет себя; однако в некоторый момент провирус или профаг может вызвать активацию вируса, который может вызвать лизис клеток-хозяев[102]. Активно размножающийся вирус не всегда убивает клетку-хозяина. Оболочечные вирусы, в том числе ВИЧ, обычно отделяются от клетки путём отпочковывания. В ходе этого процесса вирус обзаводится своей оболочкой, которая представляет собой модифицированный фрагмент клеточной мембраны хозяина или другой внутренней мембраны[103]. Таким образом, клетка может продолжать жить и продуцировать вирус. Действие на клетки
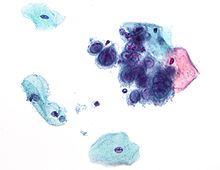
Микрофотография, показывающая цитопатические эффекты, вызванные вирусом простого герпеса первого типа. Тест Папаниколау Диапазон структурных и биохимических эффектов, оказываемых вирусом на инфицированную клетку, очень широк[109]. Они называются цитопатическими эффектами [110]. Большинство вирусных инфекций приводят к гибели клеток-хозяев. Причинами гибели могут быть лизис клетки, изменения клеточной мембраны и апоптоз[111]. Часто причиной гибели клетки является подавление её нормальной активности белками вируса, не все из которых входят в состав вирусной частицы[112]. Некоторые вирусы не вызывают никаких видимых изменений в поражённой клетке. Клетки, в которых вирус находится в латентном состоянии и неактивен, имеют мало признаков инфекции и нормально функционируют[113]. Это является причиной хронических инфекций, и вирус при них может никак себя не проявлять многие месяцы или годы. Так часто бывает, например, с вирусом герпеса[114][115]. Некоторые вирусы, например вирус Эпштейна — Барр, могут вызывать быстрое размножение клеток без появления злокачественности[116], в то время как другие, такие как папилломавирусы, могут вызвать рак[117]. Диапазон хозяев Вирусы, несомненно, являются самыми многочисленными биологическими объектами на Земле, и по этому показателю они превосходят все организмы, вместе взятые[118]. Они поражают все формы клеточных организмов, включая животных, растения, бактерии и грибы[7]. Тем не менее различные типы вирусов могут поражать только ограниченный круг хозяев, многие вирусы видоспецифичны. Некоторые, как, например, вирус оспы, могут поражать только один вид — людей[119], в подобных случаях говорят, что вирус имеет узкий диапазон хозяев. Напротив, вирус бешенства может поражать различные виды млекопитающих, то есть он имеет широкий диапазон хозяев[120]. Вирусы растений безвредны для животных, а большинство вирусов животных безопасны для человека[121]. Диапазон хозяев некоторых бактериофагов ограничивается одним штаммом бактерий, и они могут использоваться для определения штаммов, вызывающих вспышки инфекционных заболеваний методом фагового типирования[122]. Распространение Вирусы распространяются многими способами: вирусы растений часто передаются от растения к растению насекомыми, питающимися растительными соками, к примеру, тлями; вирусы животных могут распространяться кровососущими насекомыми, такие организмы известны как переносчики. Вирус гриппа распространяется воздушно-капельным путём при кашле и чихании. Норовирус и ротавирус, обычно вызывающие вирусные гастроэнтериты, передаются фекально-оральным путём при контакте с заражённой пищей или водой. ВИЧ является одним из нескольких вирусов, передающихся половым путём и при переливании заражённой крови. Каждый вирус имеет определённую специфичность к хозяевам, определяющуюся типами клеток, которые он может инфицировать. Круг хозяев может быть узок или, если вирус поражает многие виды, широк[123]. Эволюция Основная статья: Эволюция вирусов Классификация В таксономии живой природы вирусы выделяют в отдельный таксон, образующий в классификации Systema Naturae 2000 вместе с доменами Bacteria, Archaea и Eukaryota корневой таксон Biota [124]. В течение XX века в систематике выдвигались предложения о создании выделенного таксона для неклеточных форм жизни (Aphanobionta Novak, 1930[125]; надцарство Acytota Jeffrey, 1971[126]; Acellularia [127]), однако такие предложения не были кодифицированы[128].
|
||||||||||||
|
|
Последнее изменение этой страницы: 2021-04-04; просмотров: 199; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.220.137.164 (0.061 с.) |
||||||||||||

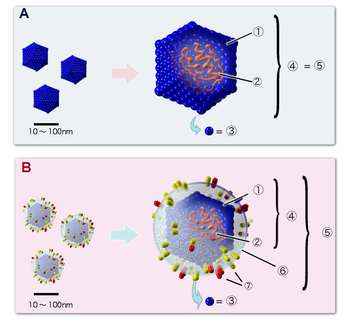 Строение икосаэдрических вирионов: А. Вирус, не имеющий липидной оболочки (например, пикорнавирус). B. Оболочечный вирус (например, герпесвирус). Цифрами обозначены: (1) капсид, (2) геномная нуклеиновая кислота, (3) капсомер, (4) нуклеокапсид, (5) вирион, (6) липидная оболочка, (7) мембранные белки оболочки
Строение икосаэдрических вирионов: А. Вирус, не имеющий липидной оболочки (например, пикорнавирус). B. Оболочечный вирус (например, герпесвирус). Цифрами обозначены: (1) капсид, (2) геномная нуклеиновая кислота, (3) капсомер, (4) нуклеокапсид, (5) вирион, (6) липидная оболочка, (7) мембранные белки оболочки
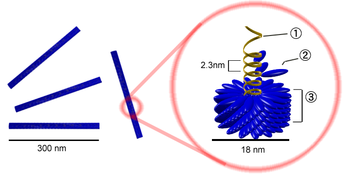 Палочковидный вирион вируса табачной мозаики. Цифрами обозначены: (1) РНК-геном вируса, (2) капсомер, состоящий всего из одного протомера, (3) зрелый участок капсида
Палочковидный вирион вируса табачной мозаики. Цифрами обозначены: (1) РНК-геном вируса, (2) капсомер, состоящий всего из одного протомера, (3) зрелый участок капсида
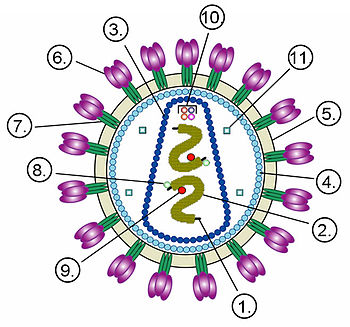 Строение вириона неикосаэдрического оболочечного вируса на примере ВИЧ.Цифрами обозначены: (1) РНК-геном вируса, (2) нуклеокапсид, (3) капсид, (4) белковый матрикс, подстилающий (5) липидную мембрану, (6) gp120 — гликопротеин, с помощью которого происходит связывание вируса с клеточной мембраной, (7) gp41 — гликопротеин. Цифрами 8—11 обозначены белки, входящие в состав вириона и необходимые вирусу на ранних стадиях инфекции: (8) — интеграза, (9) — обратная транскриптаза, (10) — Vif, Vpr, Nef и p7, (11) — протеаза
Строение вириона неикосаэдрического оболочечного вируса на примере ВИЧ.Цифрами обозначены: (1) РНК-геном вируса, (2) нуклеокапсид, (3) капсид, (4) белковый матрикс, подстилающий (5) липидную мембрану, (6) gp120 — гликопротеин, с помощью которого происходит связывание вируса с клеточной мембраной, (7) gp41 — гликопротеин. Цифрами 8—11 обозначены белки, входящие в состав вириона и необходимые вирусу на ранних стадиях инфекции: (8) — интеграза, (9) — обратная транскриптаза, (10) — Vif, Vpr, Nef и p7, (11) — протеаза