Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Основні теоретичні відомості про ергостеринСодержание книги Поиск на нашем сайте
ЗМІСТ ВСТУП 1. ОСНОВНІ ТЕОРЕТИЧНІ ВІДОМОСТІ ПРО ЕРГОСТЕРИН 1.1 Ергостерин 1.2 Продуценти ергостерину 1.3 Біосинтез ергостерину 1.4 Умови утворення ергостерину дріжджами 2. ТЕХНОЛОГІЯ БІОСИНТЕЗУ ЕРГОСТЕРИНУ 2.1 Технологічні аспекти отримання ергостерину 2.2 Накопичення біомаси дріжджів 2.3 Виробництво концентрату вітамінів групи В 2.4 Виробництво концентрату провітаміна D2 та технологія його трансформації у вітамін D2 3. РОЗРАХУНОК ФЕРМЕНТЕРА МАРКИ Б-50 3.1 Розрахунок габаритних розмірів ферментера 3.2 Розрахунок товщини стінки ферментера ВИСНОВКИ СПИСОК ВИКОРИСТАНОЇ ЛІТЕРАТУРИ
Вступ У 1912 р. для позначення додаткових харчових факторів, що у малих кількостях ефективні для лікування ряду захворювань, К. Функ ввів термін вітамін. Тоді виділяли лише 2 вітаміни — А (жиророзчинний), В (водорозчинний), сьогодні їх кількість сягає десятків. Вітаміни - це біологічно активні низькомолекулярні природні, органічні сполуки, різної хімічної структури, учасники обміну речовин і беруть участь в перетворенні енергії, переважно як компоненти ферментів. Існування і значення вітамінів відкрив російський лікар М. Лунін у кінці ХІХ ст.. Польський хімік К. Функ назвав біологічно активну речовину вітаміном, бо вона містила у своїй молекулі аміногрупу. Вітаміни мають надзвичайно велике значення для нормального обміну речовин і життєдіяльності, перебуваючи у дуже малих кількостях в продуктах харчування. За нормального раціону і зорового способу життя потреба у вітамінах задовольняється природним шляхом. За нестачі вітамінів групи D, у дітей переважно перших трьох років життя з'являються ознаки рахіту. В дорослих (особливо у вагітних жінок), які мало перебувають на сонці, не вживають достатньо повноцінної їжі, кісткова тканина втрачає кальцій і кістки розм'якшуються. В цих випадках таке явище називають остеопорозом. Недостатність вітамінів групи D може розвинутись і у дітей старших трьох років, особливо в періоди інтенсивного росту, якщо їх білкове харчування є недостатнім, а також має місце різка зміна кліматичних умов. Крім того, до розвитку D-вітамінної недостатності ведуть хронічна ниркова недостатність, хвороби печінки, тривалий прийом протисудомних препаратів, синдром мальабсорбції (порушеного всмоктування в кишечнику) різного генезу. Тому важливе значення має пошук нових безпечних джерел надходження вітамінів у організм. У випадку із вітаміном D2, використання препаратів мікробіологічного синтезу має значні переваги перед синтетичними аналогами, і з точки зору сприйнятливості організмом, і з точки зору економії енергоресурсів, затрачених на процес синтезу. Використання провітаміну D2 (ергостерину) також є більш доцільним, аніж використання готового препарату D2, оскільки організм людини здатний самостійно перетворювати провітамін у вітамінну форму, за умови достатньої кількості субстрату та сонячного світла. На сьогоднішній день перспективним є пошук нових, більш ефективних продуцентів, а також нових технологічних схем виділення та очистки кінцевого продукту.
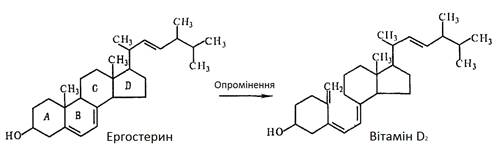
Основні теоретичні відомості про ергостерин Ергостерин Природні джерела вітаміну D досить обмежені. Відомо, що вітамін D зустрічається лише в деяких тваринних та рибних продуктах (риб’ячий жир, печінка риб, вершкове масло, молоко, ікра), а також в яєчному жовтку. Рослини, як правило, не містять вітамінів групи D, проте серед стеринів рослинних жирів виявлено провітамін D – ергостерин. Багаті на вітамін D лише печінкові жири риб. Деякі з них, наприклад печінковий жир тунця, містить в 1 г від 40 до 60 од. вітаміну D2. В молоці, вершковому маслі і яйцях вміст вітаміну D2 обмежений. Наприклад, в 100 г молока міститься лише 1 од. D2 , у вершковому маслі – 40 – 320 од. Незначний вміст вітаміну D2 в природніх продуктах змушує шукати багаті природні джерела провітаміну D2 – ергостерину. В 30-х роках ХХ ст. рядом дослідників було виявлено, що хлібопекарські дріжджі являються багатим джерелом ергостерину. Пізніше було встановлено, що деякі мікроорганізми можуть слугувати джерелом вітаміну D [2]. Ергостерин – ергоста-5,7,22-трієн-3β-ол – вихідний продукт виробництва жиророзчинного вітаміну D2 і кормових препаратів, збагачених вітаміном D2. В групу вітамінів D об’єднують споріднені сполуки, найважливішими з яких є вітаміни D2 і D3, що мають антирахітичну дію. Вітамін D2 (ергокальциферол) утворюється під час опромінення ультрафіолетовим випромінюванням ергостерину, вітамін D3 (холекальциферол) утворюється із 7-дегідрохолестерину. В організмі людини і тварин ці сполуки регулюють засвоєння кальцію та фосфору із їжі і відкладання їх в кістковій тканині [1].
Для прикладної медицини найбільше значення мають вітаміни D2 та D3. Нестача вітаміну D2 погіршує абсорбцію кальцію в кишківнику, у зв’язку з чим його вміст в крові також зменшується. Це викликає підвищену активність паращитовидних залоз, завдяки чому рівень кальцію в крові утримується на невисокому рівні за рахунок мобілізації його із запасів організму. Одночасно спостерігається підвищене виділення нирками фосфору, оскільки гіперпродукція паратиреоїдного гормону знижує реабсорбція фосфору в ниркових каналах. Таким чином вітамін D2 відграє важливу роль в забезпеченні організму кальцієм, перш за все, за рахунок регуляції його засвоєння з їжі. Загальновідомо, що нестача вітаміну D2 в організмі відбувається порушення мінералізації в процесі кісткоутворення, що призводить до серйозних змін в кістковому скелеті. В основі структури ергостерину і вітаміну D лежать чотири вуглецевих цикла (A, B, C, D). В молекулі вітаміну D кільце В розімкнуте. Вуглеводнева структура ергостерину та вітаміну D визначають їхні ліпофільні властивості [1].
Продуценти ергостерину Джерелом ергостерину є фітопланктон, бурі і зелені водорості, але особливо багаті на ергостерин дріжджі та плісняві гриби, які слугують сировиною для його промислового виробництва. Ергостерин – основний стерин дріжджів, на який припадає 60 – 90 % від інших стеринів: вміст ергостерину складає 0,2 – 0,5 %, але в деяких випадках досягає 10 % від сухої біомаси. Культурні раси дріжджів завжди містять більше стеринів, ніж дикі. Найбільшу кількість стеринів містять пекарські та пивні дріжджі. У відношенні ергостеролсинтезуючої здатності (% ергостеролу в абсолютно сухих дріжджах) дріжджі під час поверхневого культивування розташовуються в наступному порядку: Saccharomyces carlsbergensis (0,49 – 4,3), S. ellipsoideus (1,2 – 1,5), Rhodotorula glutinis (0,7 – 0,9), Candida utilis (0,4 – 0,6), C. tropicalis (0,2 – 0,3). В міцелії грибів Aspergillus і Penicillium вміст стеринів може досягати 1,2 – 1,4 % (P. westlingii близько 2,2 %) в розрахунку на сухий міцелій. Бактерії, як правило, синтезують дуже малі кількості стеринів. Зазвичай, вміст стеринів в їхніх клітинах складає 0,001 – 0,1 мг/г сухої біомаси. Стерини виявлено в Lactobacillus arabinosus, L. pentosus, Escherichia coli, Azotobacter chroococcum, Micromonospora sp., Streptomyces griseus, Sphaerotillis natans, Rhodospirillum rubrum. Але відомо два представника бактерій: Halobacterium cutirubrum і Methylococcus capsulatus, що синтезують високі кількості сквалену (1,0 і 5,5 мг відповідно на грам сухих клітин). Сквален і його чотири гідроформи виділені із метанутворюючої бактерії Methanobacillus kuzneceovii [1].
Біосинтез ергостерину Стерини, каротиноїди, сполуки груп Q-коферментів належать до терпенів і мають спільний шлях біосинтезу за «ізопреновим правилом». У відповідності до цього правила, каротиноїди (політерпени), стерини (тритерпени), а також убіхінони і гіберелінова кислота синтезуються із ізопренових одиниць в результаті проходження чотирьох стадій: 1) утворення мевалонату із ацетил-КоА або лейцину; 2) дегідратація і декарбоксилювання мевалонілпірофосфату з утворенням «активного ізопрену» - ізопентенілпірофосфату і конденсація ізопренових ланок з утворенням ациклічних терпенів різної довжини; 3) циклювання ациклічних структур; 4) подальша модифікація циклічної структури. Інтермедіатами синтезу стеринів є ацетат, мевалонова кислота, сквален, ланостерин. Сквален – загальний попередник стеринів рослинного і тваринного походження, накопичується в дріжджах і під час аерування перетворюється в стерин. Стартовою сполукою в біосинтезі стеринів є ацетат. Дві молекули ацетил-КоА конденсуються із утворенням ацетоацетил-КоА, який в свою чергу конденсується з ще одною молекулою ацетил-КоА, утворюючи β-гідрокси-β-метилглутарил-КоА. Під час відновлення даної сполуки утворюється мевалонова кислота (МВК), котра за наявності АТФ фосфорилюється із утворенням пірофосфату МВК. За наявності АТФ шляхом декарбоксилювання і дегідрування пірофосфат МВК перетворюється в 5-вуглеводневу ізопренову одиницю – ізопентенілпірофосфат. Ізопентенілпірофосфат (ІПФ) ізомерується до стадії деметилалілпірофосфату (ДМАПФ). Потім відбувається конденсація ІПФ і ДМАПФ з утворенням геранілпірофосфату. Ці сполуки, що містять 10 атомів вуглецю, конденсуються з ІПФ з утворенням фарнезілпірофосфату
Фарнезілпірофосфат далі димеризується із утворенням сквалену С15 + С15 = С30:
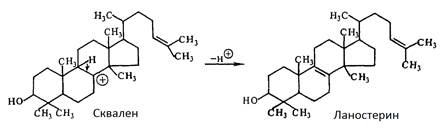
Під час циклювання і відщіплення протону утворюється ланостерин, попередник холестерину і ергостерину:
Перетворення ланостерину в ергостерин відбувається в результаті наступних стадій: 1) деметилювання ланостерину; 2) трансалкілування з утворенням 24(28)-метиленової групи і одночасним відновленням С-24(25) подвійного зв’язку; 3) десатурація бічного ланцюга з утворенням С-22(23) подвійного зв’язку; 4) ізомеризація ∆8∆7; 5) дегідрування з утворенням ∆5; 6) відновлення 24(28) метиленової групи до метильної [1].
Висновки 1. Процес накопичення біомаси, збагаченої провітаміном D2 (ергостерином), найкраще проводити на поживному середовищі, що містить н -алкани, як найоптимальніший субстрат для отримання цільового продукту. 2. Для отримання біомаси продуцента на середовищі, збагаченому н -алканами, рекомендовано використовувати 12-секційний тороїдний біореактор марки Б-50. Даний ферментер забезпечує специфічні особливості кінетики накопичення біомаси та умови перемішування і аерації. 3. Для вилучення вітамінів, міцно зв’язаних із білково-ферментними комплексами, і збільшення таким чином їхнього виходу, необхідно піддавати дріжджі гідролізу і автолізу. 4. Для забезпечення належної консистенції необхідно автолізовану (гідролізовану) дріжджову масу піддавати обробці спиртом з екстракцією водорозчинних вітамінів групи В. 5. Для вилучення стеринів і фосфоліпідів висушений дріжджовий шріт варто піддавати додатковій обробці спиртом або іншим розчинником. 6. Отриманий після екстракції стеринів та видалення спирту білково-дріжджовий шріт можна використовувати для харчових або кормових цілей, а також для отримання нуклеїнових кислот. 7. Для конструювання ферментера марки Б-50 продуктивністю 200 кг вологої біомаси на годину приймемо наступні габаритні розміри: - зовнішній радіус тороїда R = 4,4 м; - внутрішній радіус тороїда r = 1,3 м; - висота секції h = R – r = 4,4 – 1,3 = 3,1 м. - товщина стінки ферментера δ = 4 мм. ЗМІСТ ВСТУП 1. ОСНОВНІ ТЕОРЕТИЧНІ ВІДОМОСТІ ПРО ЕРГОСТЕРИН 1.1 Ергостерин 1.2 Продуценти ергостерину 1.3 Біосинтез ергостерину 1.4 Умови утворення ергостерину дріжджами 2. ТЕХНОЛОГІЯ БІОСИНТЕЗУ ЕРГОСТЕРИНУ 2.1 Технологічні аспекти отримання ергостерину 2.2 Накопичення біомаси дріжджів 2.3 Виробництво концентрату вітамінів групи В 2.4 Виробництво концентрату провітаміна D2 та технологія його трансформації у вітамін D2 3. РОЗРАХУНОК ФЕРМЕНТЕРА МАРКИ Б-50 3.1 Розрахунок габаритних розмірів ферментера 3.2 Розрахунок товщини стінки ферментера ВИСНОВКИ СПИСОК ВИКОРИСТАНОЇ ЛІТЕРАТУРИ
Вступ У 1912 р. для позначення додаткових харчових факторів, що у малих кількостях ефективні для лікування ряду захворювань, К. Функ ввів термін вітамін. Тоді виділяли лише 2 вітаміни — А (жиророзчинний), В (водорозчинний), сьогодні їх кількість сягає десятків. Вітаміни - це біологічно активні низькомолекулярні природні, органічні сполуки, різної хімічної структури, учасники обміну речовин і беруть участь в перетворенні енергії, переважно як компоненти ферментів. Існування і значення вітамінів відкрив російський лікар М. Лунін у кінці ХІХ ст.. Польський хімік К. Функ назвав біологічно активну речовину вітаміном, бо вона містила у своїй молекулі аміногрупу. Вітаміни мають надзвичайно велике значення для нормального обміну речовин і життєдіяльності, перебуваючи у дуже малих кількостях в продуктах харчування. За нормального раціону і зорового способу життя потреба у вітамінах задовольняється природним шляхом. За нестачі вітамінів групи D, у дітей переважно перших трьох років життя з'являються ознаки рахіту. В дорослих (особливо у вагітних жінок), які мало перебувають на сонці, не вживають достатньо повноцінної їжі, кісткова тканина втрачає кальцій і кістки розм'якшуються. В цих випадках таке явище називають остеопорозом. Недостатність вітамінів групи D може розвинутись і у дітей старших трьох років, особливо в періоди інтенсивного росту, якщо їх білкове харчування є недостатнім, а також має місце різка зміна кліматичних умов. Крім того, до розвитку D-вітамінної недостатності ведуть хронічна ниркова недостатність, хвороби печінки, тривалий прийом протисудомних препаратів, синдром мальабсорбції (порушеного всмоктування в кишечнику) різного генезу. Тому важливе значення має пошук нових безпечних джерел надходження вітамінів у організм. У випадку із вітаміном D2, використання препаратів мікробіологічного синтезу має значні переваги перед синтетичними аналогами, і з точки зору сприйнятливості організмом, і з точки зору економії енергоресурсів, затрачених на процес синтезу. Використання провітаміну D2 (ергостерину) також є більш доцільним, аніж використання готового препарату D2, оскільки організм людини здатний самостійно перетворювати провітамін у вітамінну форму, за умови достатньої кількості субстрату та сонячного світла. На сьогоднішній день перспективним є пошук нових, більш ефективних продуцентів, а також нових технологічних схем виділення та очистки кінцевого продукту.
Основні теоретичні відомості про ергостерин Ергостерин Природні джерела вітаміну D досить обмежені. Відомо, що вітамін D зустрічається лише в деяких тваринних та рибних продуктах (риб’ячий жир, печінка риб, вершкове масло, молоко, ікра), а також в яєчному жовтку. Рослини, як правило, не містять вітамінів групи D, проте серед стеринів рослинних жирів виявлено провітамін D – ергостерин. Багаті на вітамін D лише печінкові жири риб. Деякі з них, наприклад печінковий жир тунця, містить в 1 г від 40 до 60 од. вітаміну D2. В молоці, вершковому маслі і яйцях вміст вітаміну D2 обмежений. Наприклад, в 100 г молока міститься лише 1 од. D2 , у вершковому маслі – 40 – 320 од. Незначний вміст вітаміну D2 в природніх продуктах змушує шукати багаті природні джерела провітаміну D2 – ергостерину. В 30-х роках ХХ ст. рядом дослідників було виявлено, що хлібопекарські дріжджі являються багатим джерелом ергостерину. Пізніше було встановлено, що деякі мікроорганізми можуть слугувати джерелом вітаміну D [2]. Ергостерин – ергоста-5,7,22-трієн-3β-ол – вихідний продукт виробництва жиророзчинного вітаміну D2 і кормових препаратів, збагачених вітаміном D2. В групу вітамінів D об’єднують споріднені сполуки, найважливішими з яких є вітаміни D2 і D3, що мають антирахітичну дію. Вітамін D2 (ергокальциферол) утворюється під час опромінення ультрафіолетовим випромінюванням ергостерину, вітамін D3 (холекальциферол) утворюється із 7-дегідрохолестерину. В організмі людини і тварин ці сполуки регулюють засвоєння кальцію та фосфору із їжі і відкладання їх в кістковій тканині [1].
Для прикладної медицини найбільше значення мають вітаміни D2 та D3. Нестача вітаміну D2 погіршує абсорбцію кальцію в кишківнику, у зв’язку з чим його вміст в крові також зменшується. Це викликає підвищену активність паращитовидних залоз, завдяки чому рівень кальцію в крові утримується на невисокому рівні за рахунок мобілізації його із запасів організму. Одночасно спостерігається підвищене виділення нирками фосфору, оскільки гіперпродукція паратиреоїдного гормону знижує реабсорбція фосфору в ниркових каналах. Таким чином вітамін D2 відграє важливу роль в забезпеченні організму кальцієм, перш за все, за рахунок регуляції його засвоєння з їжі. Загальновідомо, що нестача вітаміну D2 в організмі відбувається порушення мінералізації в процесі кісткоутворення, що призводить до серйозних змін в кістковому скелеті. В основі структури ергостерину і вітаміну D лежать чотири вуглецевих цикла (A, B, C, D). В молекулі вітаміну D кільце В розімкнуте. Вуглеводнева структура ергостерину та вітаміну D визначають їхні ліпофільні властивості [1].
Продуценти ергостерину Джерелом ергостерину є фітопланктон, бурі і зелені водорості, але особливо багаті на ергостерин дріжджі та плісняві гриби, які слугують сировиною для його промислового виробництва. Ергостерин – основний стерин дріжджів, на який припадає 60 – 90 % від інших стеринів: вміст ергостерину складає 0,2 – 0,5 %, але в деяких випадках досягає 10 % від сухої біомаси. Культурні раси дріжджів завжди містять більше стеринів, ніж дикі. Найбільшу кількість стеринів містять пекарські та пивні дріжджі. У відношенні ергостеролсинтезуючої здатності (% ергостеролу в абсолютно сухих дріжджах) дріжджі під час поверхневого культивування розташовуються в наступному порядку: Saccharomyces carlsbergensis (0,49 – 4,3), S. ellipsoideus (1,2 – 1,5), Rhodotorula glutinis (0,7 – 0,9), Candida utilis (0,4 – 0,6), C. tropicalis (0,2 – 0,3). В міцелії грибів Aspergillus і Penicillium вміст стеринів може досягати 1,2 – 1,4 % (P. westlingii близько 2,2 %) в розрахунку на сухий міцелій. Бактерії, як правило, синтезують дуже малі кількості стеринів. Зазвичай, вміст стеринів в їхніх клітинах складає 0,001 – 0,1 мг/г сухої біомаси. Стерини виявлено в Lactobacillus arabinosus, L. pentosus, Escherichia coli, Azotobacter chroococcum, Micromonospora sp., Streptomyces griseus, Sphaerotillis natans, Rhodospirillum rubrum. Але відомо два представника бактерій: Halobacterium cutirubrum і Methylococcus capsulatus, що синтезують високі кількості сквалену (1,0 і 5,5 мг відповідно на грам сухих клітин). Сквален і його чотири гідроформи виділені із метанутворюючої бактерії Methanobacillus kuzneceovii [1].
Біосинтез ергостерину Стерини, каротиноїди, сполуки груп Q-коферментів належать до терпенів і мають спільний шлях біосинтезу за «ізопреновим правилом». У відповідності до цього правила, каротиноїди (політерпени), стерини (тритерпени), а також убіхінони і гіберелінова кислота синтезуються із ізопренових одиниць в результаті проходження чотирьох стадій: 1) утворення мевалонату із ацетил-КоА або лейцину; 2) дегідратація і декарбоксилювання мевалонілпірофосфату з утворенням «активного ізопрену» - ізопентенілпірофосфату і конденсація ізопренових ланок з утворенням ациклічних терпенів різної довжини; 3) циклювання ациклічних структур; 4) подальша модифікація циклічної структури. Інтермедіатами синтезу стеринів є ацетат, мевалонова кислота, сквален, ланостерин. Сквален – загальний попередник стеринів рослинного і тваринного походження, накопичується в дріжджах і під час аерування перетворюється в стерин. Стартовою сполукою в біосинтезі стеринів є ацетат. Дві молекули ацетил-КоА конденсуються із утворенням ацетоацетил-КоА, який в свою чергу конденсується з ще одною молекулою ацетил-КоА, утворюючи β-гідрокси-β-метилглутарил-КоА. Під час відновлення даної сполуки утворюється мевалонова кислота (МВК), котра за наявності АТФ фосфорилюється із утворенням пірофосфату МВК. За наявності АТФ шляхом декарбоксилювання і дегідрування пірофосфат МВК перетворюється в 5-вуглеводневу ізопренову одиницю – ізопентенілпірофосфат. Ізопентенілпірофосфат (ІПФ) ізомерується до стадії деметилалілпірофосфату (ДМАПФ). Потім відбувається конденсація ІПФ і ДМАПФ з утворенням геранілпірофосфату. Ці сполуки, що містять 10 атомів вуглецю, конденсуються з ІПФ з утворенням фарнезілпірофосфату
Фарнезілпірофосфат далі димеризується із утворенням сквалену С15 + С15 = С30:
Під час циклювання і відщіплення протону утворюється ланостерин, попередник холестерину і ергостерину:
Перетворення ланостерину в ергостерин відбувається в результаті наступних стадій: 1) деметилювання ланостерину; 2) трансалкілування з утворенням 24(28)-метиленової групи і одночасним відновленням С-24(25) подвійного зв’язку; 3) десатурація бічного ланцюга з утворенням С-22(23) подвійного зв’язку; 4) ізомеризація ∆8∆7; 5) дегідрування з утворенням ∆5; 6) відновлення 24(28) метиленової групи до метильної [1].
|
||||
|
|
Последнее изменение этой страницы: 2020-03-02; просмотров: 249; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.12.165.68 (0.01 с.) |