
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Кафедра биологии с экологиейСодержание книги Поиск на нашем сайте Кафедра биологии с экологией А.Б.Ходжаян Н.Н. Федоренко Л.А. Краснова К НЕКОТОРЫМ ВОПРОСАМ ЭВОЛЮЦИИ (дополненное) Методическое пособие для студентов 1-го курса СтГМА
СТАВРОПОЛЬ, УДК 57:575. К некоторым вопросам эволюции. Методическое пособие для студентов 1 курсов. Изд: СтГМА. 2009г.с.31.
В учебнике биологии под ред. А.А.Слюсарева и С.В. Жуковой, которым пользуются студенты 1-го курса при изучении медицинской биологии и генетики, некоторые вопросы теории эволюции требуют дополнения и уточнения. Сотрудники кафедры биологии СтГМА сочли необходимым составить данное методическое пособие по некоторым вопросам теории эволюции живой природы.
Составители: д.м.н., проф. Ходжаян А.Б., к.м.н., доц. Федоренко Н.Н., к.м.н., доц. Краснова Л.А. © Ставропольская государственная медицинская академия, 2009 ЭВОЛЮЦИЯ ГОЛОВНОГО МОЗГА У ПОЗВОНОЧНЫХ Формирование головного мозга у зародышей всех позвоночных начинается с появления на переднем конце нервной трубки вздутий - мозговых пузырей. Вначале их образуется три, а затем пять. Из переднего мозгового пузыря в дальнейшем образуется передний и промежуточный мозг, из среднего - средний мозг, а из заднего - мозжечок и продолговатый мозг. Последний без резкой границы переходит в спинной мозг В нервной трубке есть полость - невроцель, которая в ходе образования пяти мозговых пузырей формирует расширения - мозговые желудочки (у человека их 4).В этих участках мозга различают дно (основание) и крышу (мантия). Крыша располагается над -, а дно под желудочками. Вещество мозга неоднородно - представлено серым и белым веществом. Серое - это скопление нейронов, а белое образовано отростками нейронов, покрытыми жироподобным веществом (миелиновой оболочкой), которое придает веществу мозга белый цвет. Слой серого вещества на поверхности крыши любого отдела мозга называется корой. Большую роль в эволюции нервной системы играют органы чувств. Именно концентрация органов чувств на переднем конце тела обусловила прогрессивное развитие головного отдела нервной трубки. Полагают, что передний мозговой пузырь сформировался под влиянием обонятельного, средний - зрительного, а задний - слухового рецепторов. Рыбы Передний мозг небольшой, не разделен на полушарии, имеет только один желудочек. Его крыша не содержит нервных элементов, а образована эпителием. Нейроны сосредоточены на дне желудочка в полосатых телах и в отходящих спереди от переднего мозга обонятельных долях. По существу, передний мозг выполняет функцию обонятельного центра. Средний мозг является высшим регуляторным и интегративным центром. Он состоит из двух зрительных долей и является наиболее крупным отделом мозга. Такой тип мозга, где высшим регуляторным центром является средний мозг, называется ихтиопсидпым. Промежуточный мозг состоит из крыши (таламуса) и дна (гипоталамуса) С гипоталамусом связан гипофиз, а с таламусом - эпифиз. Мозжечок у рыб хорошо развит, поскольку их движения отличаются большим разнообразием. Продолговатый мозг без резкой границы переходит в спинной мозг и в нем сосредоточен пищевой, сосудодвигательный и дыхательный центры. От мозга отходит 10 пар черепно-мозговых нервов, что характерно для низших позвоночных. Амфибии У амфибий имеется ряд прогрессивных изменений в головном мозге, что связано с переходом к наземному образу жизни, где условия по сравнению с водной средой более разнообразны и характеризуются непостоянством действующих факторов. Это привело к прогрессивному развитию органов чувств и соответственно - прогрессивному развитию головного мозга. Передний мозг у амфибии в сравнении с рыбами значительно крупнее, в нем появилось два полушария и два желудочка. В крыше переднего мозга появились нервные волокна, образующие первичный мозговой свод - архипаллиум. Тела нейронов располагаются в глубине, окружая желудочки, в основном в полосатых телах. Все еще хорошо развиты обонятельные доли. Высшим интегративным центром остается средний мозг (ихтиопсидный тип). Строение такое же, как у рыб. Мозжечок связи с примитивностью движений амфибий имеет вид небольшой пластинки. Промежуточный и продолговатый мозг такие же, как у рыб. От головного мозга отходят 10 пар черепно-мозговых нервов.
Пресмыкающиеся (рептилии) Рептилии относятся к высшим позвоночным и характеризуются более активным образом жизни, что сочетается с прогрессивным развитием всех отделов головного мозга. Передний мозг является самым крупным отделом головного мозга. Кпереди от него отходят развитые обонятельные доли. Крыша остается тонкой, но на медиальной и латеральной стороне каждого полушария появляются островки коры. Кора имеет примитивное строение и называется древней – археокортекс. Роль высшего интегративного центра выполняют полосатые тела переднего мозга - зауропсидный типмозга. Полосатые тела обеспечивают анализ поступающей информации и выработку ответных реакций. Промежуточный, мозг, будучи связан с эпифизом и гипофизом имеет еще и дорсальный придаток - теменной орган, воспринимающий световые раздражения. Средний мозг теряет значение высшего интегративного центра, снижается и его значение как зрительного центра, в связи, с чем его размеры уменьшаются. Мозжечок развит значительно лучше, чем у амфибий. Продолговатый мозг образует резкий изгиб, характерный для высших позвоночных, в том числе и для человека. От головного мозга отходит 12 пар черепно-мозговых нервов, что типично для всех высших позвоночных, в том числе и для человека. Птицы Нервная система в связи с общим усложнением организации, приспособленностью к полету и обитанием в самых различных средах, развита значительно лучше, чем у пресмыкающихся. Дня птиц характерно дальнейшее увеличение общего объема головного мозга, особенно переднего. Передний мозгу птиц - это высший интегративный центр. Его ведущим отделом являются полосатые тела (зауропсидный тип мозга). Крыша остается слабо развитой. В ней сохраняются только медиальные островки коры, которые выполняют функцию высшего обонятельного центра. Они оттесняются к перемычке между полушариями и носят название гиппокампа. Обонятельные доли развиты слабо. Промежуточный мозг небольших размеров и связан с гипофизом и эпифизом. Средний мозг имеет хорошо развитые зрительные доли, что обусловлено ведущей ролью зрения в жизни птиц. Мозжечок крупный, имеет среднюю часть с поперечными бороздами и небольшие боковые выросты. Продолговатый мот такой же, как у рептилий. 12 пар черепно-мозговых нервов. Млекопитающие Передний мозг - это наиболее крупный отдел головного мозга. У разных видов его абсолютный и относительный размеры весьма варьируют. Главная особенность переднего мозга - значительное развитие коры полушарий, которая собирает всю сенсорную информацию от органов чувств, производит высший анализ и синтез этой информации и становится аппаратом тонкой условно-рефлекторной деятельности, а у высокоорганизованных млекопитающих - и психической деятельности (маммальныйтип мозга). У наиболее высокоорганизованных млекопитающих кора имеет борозды и извилины, что значительно увеличивает ее поверхность.
Промежуточный мозг содержит около 40 ядер. Специальные ядра таламуса перерабатывают зрительные, тактильные, вкусовые и интероцептивные сигналы, направляя их затем в соответствующие зоны коры больших полушарий. В гипоталамусе сосредоточены высшие вегетативные центры, управляющие работой внутренних органов через нервные и гуморальные механизмы. В среднем мозге на смену двухолмия приходит четверохолмие. Его передние холмы являются зрительными, а задние связаны со слуховыми рефлексами. В центре среднего мозга проходит ретикулярная формация, которая служит источником восходящих влияний, активирующих кору больших полушарий. Хотя передние доли являются зрительными, анализ зрительной информации осуществляется в зрительных зонах коры, а на долю среднего мозга приходится главным образом управление глазной мускулатурой - изменение просвета зрачка, движения глаз, напряжение аккомодации. В задних холмах расположены центры, регулирующие движения ушных раковин, натяжение барабанной перепонки, перемещение слуховых косточек. Средний мозг также участвует в регуляции тонуса скелетной мускулатуры. Мозжечок имеет развитые боковые доли (полушария), покрытые корой, и червь. Мозжечок связан со всеми отделами нервной системы, имеющими отношение к управлению движениями - с передним мозгом, стволом мозга и вестибулярным аппаратом. Он обеспечивает координацию движений. Продолговатый мозг. В нем по бокам обособляются пучки нервных волокон, идущих к мозжечку, а на нижней поверхности - продолговатые валики, получившие название пирамид. От основания мозга отходит 12 пар черепно-мозговых нервов. Эволюция артериальных дуг. У эмбрионов всех позвоночных животных впереди от сердца закладывается непарная брюшная аорта, от которой отходят артериальные дуги. Они гомологичным артериальным дугам ланцетника. Но число их меньше, чем у ланцетника: у рыб - 6-7 пар, а у наземных позвоночных 6 пар. Первые две пары у всех позвоночных испытывают редукцию. Следующие пары артериальных дуг у рыб разделяются на приносящие и выносящие жаберные артерии, а у наземных животных подвергаются сильным преобразованиям. Так, из 3-й пары дуг образуются сонные артерии. Четвертая пара преобразуется в дуги аорты, которые у амфибий и рептилии развиваются симметрично. У птиц левая дуга атрофируется и сохраняется только правая. У млекопитающих редуцируется правая дуга и сохраняется лишь левая. Пятая пара дуг у всех позвоночных редуцируется и только у хвостатых амфибий от нее сохраняется незначительный проток. Шестая дуга теряет связь со спинной аортой, от нее берут начало легочные артерии. Сосуд, связывающий во время эмбрионального развития легочную артерию со спинной аортой, называется боталловымпротоком. Во взрослом состоянии он сохраняется у хвостатых амфибий и некоторых рептилий. Как порок развития, этот проток может сохраняться и у других более высокоорганизованных животных и человека. В тесной связи с кровеносной системой находится лимфатическая система: Лимфа выполняет важную роль в обмене веществ, поскольку является посредником между кровью и тканевой жидкостью. Кроме того, она богата лейкоцитами, играющими важную роль в иммунитете. РАЗВИТИЕ СЕРДЦА
В эмбриогенезе человека наблюдается ряд филогенетических преобразований сердца, что имеет важное значение для понимания механизмов развития врожденных пороков сердца. У низших позвоночных (рыбы, амфибии) сердце закладывается под глоткой в виде полой трубки. У высших позвоночных и человека сердце закладывается в виде двух далеко отстоящих друг от друга трубок. Позже они сближаются, перемещаясь под кишку, а затем смыкаются, образуя единую трубку, расположенную посередине. У всех позвоночных передняя и задняя части трубки дают начало крупным сосудам. Средняя же часть начинает быстро и неравномерно расти, образуя S-образную форму. После этого задняя часть трубки перемещается на спинную сторону и вперед, образуя предсердие. Передняя часть трубки при этом не смещается, стенки ее утолщаются и она преобразуется в желудочек. У рыб одно предсердие, а у амфибий оно разделяется растущей перегородкой надвое. Желудочек у рыб и амфибий один, но в желудочке у амфибий имеются мышечные выросты (трабекулы)» образующие мелкие пристеночные камеры. У рептилий в желудочке образуется неполная перегородка, растущая снизу вверх. У птиц и млекопитающих желудочек разделен на две половины - правую и левую. В ходе эмбриогенеза у млекопитающих и человека вначале имеется одно предсердие и один желудочек, отделяющиеся друг от друга перехватом с каналом, сообщающим предсердие с желудочком. Затем в предсердии спереди назад начинает расти перегородка, разделяющая предсердие на две части - левое и правое. Одновременно с дорсальной и вентральной сторон начинают расти выросты, которые соединяясь, два отверстия: правое и левое. Позднее в этих отверстиях формируются клапаны. Межжелудочковая перегородка образуется из разных источников.
ЭВОЛЮЦИЯ ИММУННОЙ СИСТЕМЫ Иммунная система осуществляет защиту организма от проникновения генетически чужеродных тел: микроорганизмов, чужих клеток, инородных тел и др. Ее действие основано на способности отличать собственные структуры организма от генетически чужеродных, элиминируя последние. В эволюции формировалось три главных формы иммунного ответа: 1) фагоцитоз, или неспецифическое уничтожение генетически чужеродного материала; 2) клеточный иммунитет, основанный на специфическом распознавании его и уничтожении Т-лимфоцитами; 3) гуморальный иммунитет, осуществляемый путем трансформации В-лимфоцитов в плазматические клетки и синтез ими антител (иммуноглобулинов). В эволюции, выделяют три этапа в формировании иммунного ответа: - квазиммунное (лат. "квази" - наподобие) распознаваниеорганизмом своих и чужеродных клеток. Этот тип реакции наблюдается, начиная от кишечнополостных и кончая млекопитающими. При этом ответе не формируется иммунной памяти, то есть еще не происходит усиления иммунной реакции на повторное проникновение чужеродного материала; - п римитивный клеточный иммунитет обнаружен у кольчатых червей и иглокожих. Он обеспечивается целомоцитами – клетками вторичной полости тела, способными уничтожать чужеродный материал. На этом этапе появляется иммунологическая память; - система интегрированного клеточного и гуморального иммунитета. Для нее характерны специфические гуморальные и клеточные реакции на чужеродные тела, наличие лимфоидных органов иммунитета, образование антител. Такого типа иммунная система не характерна для беспозвоночных. Круглоротые уже способны формировать антитела, но вопрос о наличии у них вилочковой железы как центрального органа иммуногенеза является пока открытым. Впервые тимус обнаруживается у рыб. Тимус, селезенка, отдельные скопления лимфоидной ткани обнаруживаются в полном объеме начиная с амфибий. У низших позвоночных (рыбы, амфибии) вилочковая железа активно выделяет антитела, что не характерно для птиц и млекопитающих. Особенность иммунной системы иммунного ответа птиц состоит в наличии особого лимфоидного органа - фабрициевой сумки. В этом органе В-лимфоциты после антигенной стимуляции способны трансформироваться в плазматические клетки, вырабатывающие антитела. У млекопитающих органы иммунной системы разделяют на 2 типа: центральные и периферические. В центральных органах иммуногенеза созревание лимфоцитов происходит без влияния антигенов. В периферических органах иммуногенеза происходит антигензависимое Т и В - размножение и дифференцирование лимфоцитов. На ранних стадиях эмбриогенеза из желточного мешка в тимус и красный костный мозг мигрируют стволовые лимфатические клетки. После рождения источником стволовых клеток становится красный костный мозг. Периферическими лимфоидными органами являются: лимфоузлы, селезенка, миндалины, лимфоидные фолликулы кишечника. К моменту рождения они еще практически не сформированы и размножение и дифференцировка в них лимфоцитов начинается только после антигенной стимуляции мигрировавших из центральных органов иммуногенеза Т-и В-лимфоцитов.
ЭВОЛЮЦИЯ ДЫХАТЕЛЬНОЙ СИСТЕМЫ. Почти все живые организмы являются аэробами, то есть дышащими воздухом. Совокупность процессов, обеспечивающих поступление и потребление О2 и выделение СО2, называется дыханием. Функция дыхания у животных разной степени организованности обеспечивается по-разному. Наиболее простой формой дыхания является диффузия газов через стенки живой клетки (у одноклеточных) или через покровы тела (кишечнополостные; плоские, круглые и кольчатые черви). Диффузное дыхание встречается также у мелких членистоногих, имеющих тонкий хитиновый покров и относительно большую поверхность тела. С усложнением организации животных формируется специальная система органов дыхания; Так уже у некоторых водных кольчецов появляются примитивные органы дыхания - наружные жабры (эпителиальные выросты с капиллярами), При этом в дыхании участвует и кожа. У членистоногих органы дыхания имеют более сложное строение и представлены у водных форм жабрами, а у наземных и вторичноводных легкими и трахеями (у наиболее древних членистоногих, таких как скорпионы, - легкие, у пауков - и легкие и трахеи, а у насекомых, - высших членистоногих, - только трахеи). Функцию органов дыхания у низших хордовых (ланцетники) берут на себя жаберные щели, по перегородкам которых проходят жаберные артерии (100 пар). Поскольку деление артерий на капилляры в жаберных перегородках отсутствуют, общая поверхность поступления О2 невелика и окислительные процессы идут на низком уровне. Соответственно этому ланцетник ведет малоподвижный образ жизни. В связи с переходом позвоночных кактивному образу жизни в органах дыхания возникают прогрессивные изменения. Так, у рыб в жаберных лепестках, в отличие от ланцетника, появляется обильная сеть кровеносных капилляров, их дыхательная поверхность резко увеличивается, поэтому число жаберных щелей у рыб сокращается до четырех. Земноводные - первые животные, вышедшие на сушу, у которых развились органы атмосферного дыхания - легкие (из выпячивания кишечной трубки). В связи с примитивностью строения (легкие представляют собой мешки с тонкими ячеистыми стенками), количество кислорода, поступающего через легкие, удовлетворяет потребность в нем организма только на 30-40%, поэтому в дыхании принимает участие и кожа, содержащая многочисленные кровеносные капилляры (кожно-легочное дыхание). Воздухоносные пути у амфибий слабо дифференцированы. Они соединены с ротоглоткой небольшой гортано-трахейной камерой.
Млекопитающие имеют легкие альвеолярного строения, благодаря чему их поверхность в 50-100 раз больше поверхности тела. Бронхи древовидно разветвлены и заканчиваются тонкостенными бронхиолами с гроздьями альвеол, густо оплетенных кровеносными капиллярами. Хорошо развиты гортань и трахея. Таким образом, основное направление эволюции дыхательной системы заключается в увеличении дыхательной поверхности, усложнении строения воздухоносных, путей и их обособлении от респираторных.
ЭВОЛЮЦИЯ ИММУННОЙ СИСТЕМЫ Иммунная система осуществляет защиту организма от проникновения в организм генетически чужеродных тел: микроорганизмов, вирусов, чужих клеток, инородных тел. Ее действие основано на способности отличать собственные структуры от генетически чужеродных, элиминируя их. В эволюции сформировалось три главных формы иммунного ответа: 1) фагоцитоз - или неспецифическое уничтожение генетически чужеродного материала; 2) клеточный иммунитет, основанный на специфическом распознавании и уничтожении такого материала Т-лимфоцитами; 3) гуморальный иммунитет, осуществляемый путем образования потомками В-лимфоцитов, так называемыми, плазматическими клетками иммуноглобулинов и связывания ими чужеродных антигенов. В эволюции выделяют три этапа формирования иммунного ответа: I этап - квазииммунное (лат. квази - наподобие, как бы) распознавание организмом своих и чужеродных клеток. Этот тип реакции наблюдается от кишечнополостных до млекопитающих. Эта реакция не связана с выработкой иммунных тел, и при этом не формируется иммунной памяти, то есть еще не происходит усиления иммунной реакции на повторное проникновение чужеродного материала. II этап - примитивный клеточный иммунитет обнаружен у кольчатых червей и иглокожих. Он обеспечивается целомоцитами - клетками вторичной полости тела, способными уничтожать чужеродный материал. На этом этапе появляется иммунологическая память. III этап - система интегрированного клеточного и гуморальго иммунитета. Для нее характерны специфические гуморальные и клеточные реакции на чужеродные тела. Характерно наличие лимфоидных органов иммунитета, образование антител. Такого типа иммунная система не характерна для беспозвоночных. Круглоротые способны формировать антитела, но вопрос о наличии у них вилочковой железы, как центрального органа иммуногенеза, является пока открытым. Впервые тимус обнаруживается у рыб. Эволюционные предшественники лимфоидных органов млекопитающих - тимус, селезенка, скопления лимфоидной ткани обнаруживаются в полном объеме у амфибий. У низших позвоночных (рыбы, амфибии) вилочковая железа активно выделяет антитела, что не характерно для птиц и млекопитающих. Особенность системы иммунного ответа птиц состоит в наличии особого лимфоидного органа - фабрициевой сумки. В этом органе образуются В-лимфоциты, которые после антигенной стимуляции способны трансформироваться в плазматические клетки и вырабатывать антитела. У млекопитающих органы иммунной системы разделяют на два типа: центральные и периферические. В центральных органах созревание лимфоцитов происходит без существенного влияния антигенов. Развитие периферических органов, наоборот, непосредственно зависит от антигенного воздействия - лишь при контакте с антигеном в них начинаются процессы размножения и дифференциации лимфоцитов. Центральным органом иммуногенеза у млекопитающих является тимус, где образуются Т-лимфоциты, а также красный костный мозг, где образуются В-лимфоциты. На ранних стадиях эмбриогенеза из желточного мешка в тимус и красный костный мозг мигрируют стволовые лимфотические клетки. После рождения источником стволовых клеток становится красный костный мозг. Периферическими лимфоидными органами являются: лимфоузлы, селезенка, миндалины, лимфоидные фолликулы кишечника. К моменту рождения они еще не сформированы практически и образование в них лимфоцитов начинается только после антигенной стимуляции, после того, как они заселяются Т-и В-лимфоцитами из центральных органов иммуногенеза.
ФИЛОГЕНЕЗ ВИСЦЕРИАЛЬНОГО ЧЕРЕПА У ПОЗВОНОЧНЫХ ЖИВОТНЫХ. Череп позвоночных состоит из двух основных отделов - осевого и висцерального. 1. Осевой - черепная коробка (мозговой череп - neurocranium) - продолжение осевого скелета, служит для защиты головного мозга и органов чувств. 2. Висцеральный - лицевой (splanchnocranium), образует опору для передней части пищеварительного тракта.
Первая дуга у большинства современных позвоночных приобретает функцию челюстного аппарата - ее называют челюстной, а вторую - тоже по функции - подъязычной или гиоидной. Остальные, начиная с третьей и до седьмой, называют жаберными, т.к. они служат опорой для жаберного аппарата. На ранних этапах развития висцеральный и осевой череп не связаны друг с другом, позднее эта связь возникает. Общие для всех зародышей позвоночных закладки семи висцеральных дуг в процессе эмбрионального развития претерпевают у представителей разных классов соответственно и различные специфические изменения.
I. Низшие рыбы (хрящевые) - Chondrichthyes 1-я, она же челюстная дуга, состоит из двух крупных хрящей, вытянутых в переднезаднем направлении: верхнего - небноквадратного - первичная верхняя челюсть, нижнего - меккелева - первичная нижняя челюсть; они сзади сращены между собой и выполняют функцию первичной челюсти. 2-я, она же подъязычная, или гиоидная дуга состоит из следующих компонентов: 1) из двух, расположенных, вверху гиомандибулярных хрящей, которые сверху соединены с черепной коробкой, снизу- с гиоидом, а впереди - с челюстной дугой - первичной верхней челюстью; 2) из двух гиоидов, расположенных ниже гиомандибулярных хрящей, которые с ними и соединены; кроме того, гиоиды соединены с первичной нижней челюстью; 3) из непарной копулы (небольшой хрящ, соединяющий оба гиоида между собой). Исходя из расположения гиомандибулярного хряща, понятно, что ему принадлежит роль подвеска, соединяющего челюстную дугу с черепом. Такой тип соединения называется гиостилией, а череп - гиостильным. Это свойственно низшим позвоночным - всем рыбам. Остальные висцеральные дуги с третьей по седьмую образуют опору для дыхательного аппарата. II.Высшие рыбы - (костные) Osteichthyes. Основное отличие касается только челюстной дуги: 1) верхний элемент челюстной дуги (верхняя челюсть) состоит вместо одного крупного небноквадратного хряща из пяти элементов - небного хряща, квадратной кости и трех крыловидных хрящей; 2) впереди от первичной верхней челюсти образуются две крупные накладные кости, снабженные крупными зубами, - эти кости и становится - вторичными верхними челюстями; 3) дистальный конец первичной нижней челюсти тоже покрывает большая зубная кость, которая выдается далеко вперед и образует вторичную нижнюю челюсть. Подъязычная дуга сохраняет прежнюю функцию, т.е. череп остается гипостильным. III.Земноводные - Amphibia. Главное отличие в новом способе соединения челюстной дуги с черепом: небный хрящ первичной верхней челюсти на всем протяжении срастается с осевым черепом, т.е. с черепной коробкой. Такой тип соединения называется аутостильный. Нижнечелюстной отдел соединяется с верхнечелюстным и тоже получает связь с черепом без подъязычной дуги. Таким образом, гиомандибулярный хрящ освобождается от функции подвеска, существенно редуцируется и приобретает новую функцию - входит в состав воздухоносной полости среднего уха в виде слуховой косточки - столбика. Часть подъязычной дуги (гиоидный хрящ), жаберные дуги образуют частичную опору для языка и подъязычный аппарат, частично гортанные хрящи, частично редуцируются. IV.Пресмыкающиеся - Reptilia. Череп аутостильный, но при этом небный хрящ первичной челюсти редуцируется и в причленении верхней челюсти к черепу участвует только квадратная кость, с ней соединяется нижняя челюсть и таким образом присоединяется к черепу. Остальная часть висцерального скелета образует подъязычный аппарат, который состоит из тела подъязычной кости и трех пар отростков. V.Млекопитающие - Mammalia. Появляется совершенно новый способ соединения с черепом нижней челюсти, которая присоединяется к нему непосредственно, образуя сустав с чешуйчатой костью черепа, что позволяет не только захватывать пищу, но и совершать сложные жевательные движения. В образовании сустава участвует только вторичная нижняя челюсть. Следовательно, квадратная кость первичной верхней челюсти теряет свою функцию подвеска и превращается в слуховую косточку - наковальню. Первичная нижняя челюсть в процессе эмбрионального развития полностью выходит из состава нижней челюсти и преобразуется в следующую слуховую косточку - молоточек. Верхний отдел подъязычной дуги, гомолог гиомандибулярного хряща преобразуется в стремечко. Все три слуховые косточки составляют единую функциональную цепь. 1-я – жаберная дуга (1-я висцеральная) и копула дают, начало телу подъязычной кости и ее задними рожкам. 2-я и 3-я жаберные дуги (4-я и 5-я висцеральные) дают начало щитовидному хрящу, который впервые появляется у млекопитающих. 4-я и 5-я жаберные дуги (1-я и 7-я висцеральные) дают материал для остальных гортанных хрящей, и возможно, для трахеальных. ЭВОЛЮЦИЯ ЗУБНОЙ СИСТЕМЫ И РОТОВЫХ ЖЕЛЕЗ ПОЗВОНОЧНЫХ Рыбы-Piscеs Зубная система - гомодонтная (зубы одинаковые). Зубы имеют коническую форму, обращены назад, служат для удержания пищи, располагаются по краю черепа, у некоторых на всей поверхности ротовой полости. Слюнных желез в ротовой полости нет, т.к. они пищу заглатывают с водой. Язык примитивный, в виде двойной складки слизистой оболочки. Крыша ротовой полости образована основанием мозгового черепа - первичное твердое небо. Ротовое отверстие окружено кожными складками - губами, которые неподвижны. Общая ротоглоточная полость. Плакоидная чешуя хрящевых рыб представляет собой пластинку с положенным на ней шипом. Пластинка лежит в кориуме вершина шипа через эпидермис выдается наружу. Вся чешуя состоит из дентина, образованного клетками кориума, вершина шипа покрыта эмалью, образуемой клетками базального слоя эпидермиса. Более крупные и усложненные плакоидные чешуи располагаются в челюстях, образуя зубы. В сущности, зубы всех позвоночных - это измененные плакоидные чешуи предков. Земноводные - Amphibia. Зубная система гомодонтная. Зубы у ряда земноводных располагаются не только на альвеолярной дуге им, как и рыбам, свойствен полифиодонтизм. Появляются слюнные железы, секрет которых не содержит ферментов. В языке располагаются мышцы, определяющие его собственную подвижность. Крыша ротовой полости - тоже первичное твердое небо. Губы неподвижны Общая ротоглоточная полость. Пресмыкающиеся - Reptilia. Зубная система у современных рептилий гомодонтная, ядовитые пресмыкающиеся имеют специальные зубы, по которым яд стекает в ранку от укуса. Зубы располагаются в один ряд. У некоторых вымерших форм обнаруживается начальная дифференцировка. Всем рептилиям свойствен полифиодонтизм. Слюнные железы развиты лучше, среди них выделяют подъязычные, зубные и губные. Секрет желез уже содержит ферменты. У ядовитых змей задняя пара зубных желез преобразована в ядовитые, секрет содержит токсины (яд). Язык закладывается из трех зачатков: один - непарный и два - парных, лежащих впереди непарного. Парные зачатки позже срастастаются. У большинства рептилий это срастание неполное и язык раздвоен. Возникают зачатки вторичного твердого неба в виде горизонтальных костных складок верхней челюсти, которые доходят до середины и делят ротовую полость на верхний отдел - дыхательный (носоглоточный) и нижний - вторичную ротовую полость. Губы неподвижны. Млекопитающие - Mammalia, Зубы гетеродонтные, т.е. дифференцированные: различают резцы (incisivi), клыки (canini), малые коренные (praemolares) и коренные (molares). У ластоногих и зубатых китов зубы не дифференцированы. Зубы сидят в альвеолах, на альвеолярных дугах челюстей основание зуба сужается, образуя корень. Резцы и клыки весьма сходны с коническими зубами предков (рептилий), коренные зубы подверглись наибольшим эволюционным преобразованиям и впервые возникли у зверозубых ящеров.
Общее количество зубов у разных отрядов – разное: например, у слонов - 6, волка - 42, кошек - 30, зайцев - 28, у большинства приматов и человека-32. Слюнные железы млекопитающих многочисленны: это и мелкие - язычные, щечные, небные, зубные - гомологичные железам рептилий, и крупные - подъязычные, подчелюстные, околоушные. Из них первые две появились в результате дифференцировки подъязычной железы рептилий, а околоушные - новое приобретение млекопитающих. В ротовой полости - у высших млекопитающих появляются крупные скопления лимфатической ткани - миндалины. Язык также, как и у пресмыкающихся, развивается из трех зачатков. Вторичное твердое небо становится сплошным, полностью отделяется полость рта от полости носа, чем достигается независимость функций ротовой полости и дыхания. Кзади твердое небо продолжается в мягкое - двойная складка слизистой, отделяющая ротовую полость глотки. Поперечные валики твердого неба способствуют перетиранию пищи. У человека они постепенно исчезают после рождения. Губы мясистые у сумчатых и плацентарных, подвижны, что связано со вскармливанием детенышей молоком. Губы, щеки и челюсти ограничивают пространство, называемое преддверием рта. У человека зубная формула 2123 2123(половина верхней и нижней челюсти). Зубы, по сравнению с другими приматами, уменьшились в размерах, особенно клыки, они не выступают из зубного ряда и не перекрываются. Исчезли диастемы (промежутки между зубами) в верхних и н
|
||
|
|
Последнее изменение этой страницы: 2016-04-07; просмотров: 588; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.62 (0.015 с.) |


 Для переднего мозга млекопитающих и человека характерна функциональная асимметрия. У человека, она выражается в том, что правое полушарие отвечает за образное мышление, и левое - за абстрактное. Кроме того, в левом полушарии находятся центры устной и письменной речи.
Для переднего мозга млекопитающих и человека характерна функциональная асимметрия. У человека, она выражается в том, что правое полушарие отвечает за образное мышление, и левое - за абстрактное. Кроме того, в левом полушарии находятся центры устной и письменной речи.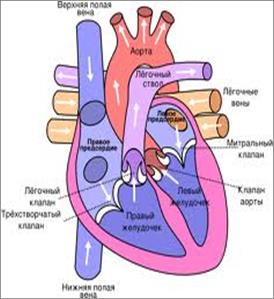 Нарушение эмбриогенеза сердца может выражаться в отсутствии или неполном заращении межпредсердной или межжелудочковой перегородки. Из аномалий развития сосудов наиболее часто встречается незаращение боталлова протока (от 6 до 22% от всех врожденных пороков сердечно-сосудистой системы), реже - незаращение сонного протока. Кроме того, может вместо одной дуги аорты развиться две - левая и правая, которые образуют аортальное кольцо вокруг трахеи и пищевода с возрастом это кольцо может сужаться и нарушаться глотание. Иногда встречается транспозиция аорты, когда она начинается не от левого желудочка, а от правого, а легочная артерия - от левого.
Нарушение эмбриогенеза сердца может выражаться в отсутствии или неполном заращении межпредсердной или межжелудочковой перегородки. Из аномалий развития сосудов наиболее часто встречается незаращение боталлова протока (от 6 до 22% от всех врожденных пороков сердечно-сосудистой системы), реже - незаращение сонного протока. Кроме того, может вместо одной дуги аорты развиться две - левая и правая, которые образуют аортальное кольцо вокруг трахеи и пищевода с возрастом это кольцо может сужаться и нарушаться глотание. Иногда встречается транспозиция аорты, когда она начинается не от левого желудочка, а от правого, а легочная артерия - от левого. У рептилий в связи с окончательным выходом на сушу происходит дальнейшее усложнение дыхательной системы: Кожное дыхание исчезает, а дыхательная поверхность легочных мешков увеличивается, благодаря появлению большого количества разветвленных перегородок, в которых проходят кровеносные капилляры. Усложняются и воздухоносные пути: в трахее формируются хрящевые кольца, разделяясь, она дает два бронха. Начинается формирование внутрилегочных бронхов.
У рептилий в связи с окончательным выходом на сушу происходит дальнейшее усложнение дыхательной системы: Кожное дыхание исчезает, а дыхательная поверхность легочных мешков увеличивается, благодаря появлению большого количества разветвленных перегородок, в которых проходят кровеносные капилляры. Усложняются и воздухоносные пути: в трахее формируются хрящевые кольца, разделяясь, она дает два бронха. Начинается формирование внутрилегочных бронхов.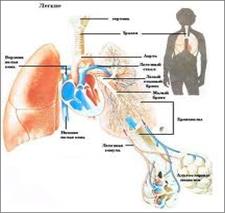 У птиц в строении органов дыхания появляется ряд особенностей. Легкие у них имеют многочисленные перегородки с сетью кровеносных капилляров. От трахеи идет бронхиальное дерево, заканчивающееся бронхиолами. Часть главных и вторичных бронхов выходит за пределы легких и образует шейные, грудные и брюшные пары воздушных мешков, а также проникает в кости, делая их пневматичными. Во время полета кровь насыщается кислородом и на акте вдоха и на акте выдоха (двойное дыхание).
У птиц в строении органов дыхания появляется ряд особенностей. Легкие у них имеют многочисленные перегородки с сетью кровеносных капилляров. От трахеи идет бронхиальное дерево, заканчивающееся бронхиолами. Часть главных и вторичных бронхов выходит за пределы легких и образует шейные, грудные и брюшные пары воздушных мешков, а также проникает в кости, делая их пневматичными. Во время полета кровь насыщается кислородом и на акте вдоха и на акте выдоха (двойное дыхание). Оба отдела черепа развиваются независимо друг от друга, разными путями. Висцеральная часть черепа у зародышей позвоночных животных состоит из метамерно расположенных хрящевых дуг, которые охватывают переднюю часть пищеварительного тракта, и отделены друг от друга висцеральными щелями. Дуги обозначают порядковыми номерами в соответствии с расположением по отношению к черепу.
Оба отдела черепа развиваются независимо друг от друга, разными путями. Висцеральная часть черепа у зародышей позвоночных животных состоит из метамерно расположенных хрящевых дуг, которые охватывают переднюю часть пищеварительного тракта, и отделены друг от друга висцеральными щелями. Дуги обозначают порядковыми номерами в соответствии с расположением по отношению к черепу. В связи с дифференцировкой зубов увеличивается продолжительность функционирования. В онтогенезе существует две смены зубов (дифиодонтизм): резцы, клыки и большие коренные имеют две генерации (молочные и постоянные); малые коренные - только одну.
В связи с дифференцировкой зубов увеличивается продолжительность функционирования. В онтогенезе существует две смены зубов (дифиодонтизм): резцы, клыки и большие коренные имеют две генерации (молочные и постоянные); малые коренные - только одну.


