
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Все без исключения растительные и животные организмы состоят из клеток.Стр 1 из 9Следующая ⇒
ГЛАВА 2. БИОЛОГИЯ КЛЕТКИ 2.1. Клетка - элементарная структурно-функциональная Клетка представляет собой наименьшую обособленную живую систему, которой присущи все свойства жизни и которая может в определённых условиях среды их сохранять и передавать в ряду поколений. Клетка несёт полную характеристику жизни. Вне клетки на планете Земля не существует полноценной жизнедеятельности. Поэтому в природе Земли клетке принадлежит роль элементарной структурной, функциональной и генетической единицы жизни. Это означает, что клетка составляет основу строения, жизнедеятельности и развития всех живых форм - одноклеточных, многоклеточных и даже неклеточных. Благодаря своим биологическим механизмам клетка осуществляет обмен веществ, использование биологической информации, размножение, реализует свойства наследственности и изменчивости, обусловливая тем самым присущие органическому миру качества единства и разнообразия. Занимая в мире живых существ положение элементарной единицы, клетка, тем не менее, отличается сложным строением. При этом определённые общие черты обнаруживаются во всех без исключения клетках, характеризуя наиболее важные стороны клеточной организации и жизнедеятельности. 2.2. Основные этапы развития Клеточная теория - обобщённое представление о строении клеток как единиц живого, об их воспроизведении и роли в формировании многоклеточных организмов. Появлению и формированию отдельных положений клеточной теории предшествовал длительный (более 300 лет) период накопления знаний о строении различных одноклеточных и многоклеточных организмов, растений и животных. Этот период связан с конструированием, применением и усовершенствованием различных светооптических приборов, в т.ч. микроскопов. Несмотря на то, что наука о клетке (цитология, биология клетки) возникла с формулированием первого крупнейшего обобщения в биоло- Первые микроскопы были созданы на рубеже ХVI и XVII веков практически одновременно в трёх странах: в Голландии микроскоп сконструировали братья Янсен (1590), в Италии - Г. Галилей (1610), в Германии - Кеплер (1617).
Значительное усовершенствование методов микроскопического исследования (создание голландскими и российскими физиками ахроматических микроскопов) произошло в начале XIX века. Это позволило Р. Броуну (1931-1933) обнаружить в растительных клетках самый крупный органоид - ядро. Позднее ядро было описано в клетках
Гиалоплазма Гиалоплазма (матрикс цитоплазмы, истинная внутренняя среда цитоплазмы) представляет собой многофазную дисперсную систему, в которой дисперсной средой является вода, а дисперсной фазой - молекулы органических и неорганических веществ (таблица), а также фрагменты биологических мембран и органоидов клетки, поглощённые клеткой частицы и вещества. Содержание в цитоплазме химических соединений
Поскольку диаметр белковых молекул превышает 0,001 мкм, а другие частицы дисперсной системы имеют размеры от 0,1 до 0,001 мкм, т.е. слишком велики, чтобы образовать истинный раствор, но и слишком малы, чтобы выпасть в осадок, возникает коллоидный раствор. Удельный вес цитоплазмы составляет в среднем 1,03. В упорядоченной многокомпонентной системе гиалоплазмы отдельные зоны могут менять своё агрегатное состояние в зависимости от условий или от функциональной задачи; в бесструктурной, на взгляд, гиалоплазме могут возникать и распадаться различные фибриллярные и нитчатые комплексы белковых молекул. В структуру гиалоплазмы входят главным образом различные глобулярные белки, составляющие 20-25% общего содержания белков эукариотической клетки. К важнейшим ферментам гиалоплазмы относятся ферменты метаболизма сахаров, азотистых оснований, аминокислот, липидов и др. соединений. В гиалоплазме располагаются ферменты активации аминокислот при синтезе белков, транспортные РНК. При участии рибосом и полирибосом (полисом) в гиалоплазме происходит синтез белков, необходимых для собственных клеточных нужд. Осмотические и буферные свойства клетки в значительной степени определяются составом и структурой гиалоплазмы. Несомненно, важнейшая роль гиалоплазмы заключается в объединении всех клеточных структур и обеспечении химического взаимодействия их друг с другом. Через гиалоплазму осуществляется большая часть внутриклеточных транспортных процессов: перенос аминокислот, жирных кислот, нуклеотидов и сахаров. В гиалоплазме идёт постоянный поток ионов к плазматической мембране и от неё к митохондриям, ядру и вакуолям. Гиалоплазма является основным вместилищем и зоной перемещения молекул АТФ, в ней происходит отложение запасаемых продуктов: гликогена, жировых капель и некоторых пигментов. Для цитоплазмы характерно постоянное движение ее коллоидных частиц и других компонентов. Гиалоплазма пронизана микротрубочками, филаментами и микрофиламентами, полимеризация и распад которых обеспечивают обратимые переходы участков цитоплазмы из золя (более жидкого состояния) в более вязкое состояние - гель. В цитоплазме осуществляются все процессы внутриклеточного метаболизма, кроме происходящего в ядре синтеза нуклеиновых кислот. Через плазматическую мембрану осуществляется обмен веществ между цитоплазмой и внешней средой, через ядерную оболочку - ядерно-цитоплазматический обмен. Под контролем ядра цитоплазма способна к росту и воспроизведению, при частичном удалении она полностью регенерирует. Однако цитоплазма не способна, как правило, к длительному автономному существованию - в безъядерных клетках она дегенерирует.
В животных клетках различают два слоя цитоплазмы: 1) наружный слой (эктоплазма), лишенный гранул и большинства органоидов, обладающий относительно высокой вязкостью; под плазматической мембраной в нем располагается сплетение микрофиламентов; 2) внутренний слой (эндоплазма), содержащий различные органоиды и гранулы, обладающий относительно меньшей вязкостью. В составе цитоплазмы клеток живых организмов обнаружено более 80 химических элементов.
Исходя из количественного содержания в клетке, химические элементы разделяются на три группы: 1. Макроэлементы, на долю которых приходится приблизительно 99% всей массы клетки. Четыре из них (углерод, кислород, азот и водород) составляют 98% от массы всех макроэлементов. Однако в эту группу включены также химические элементы, содержание которых в клетке достигает 0,1 - 0,01% (калий, магний, кальций, железо, сера, фосфор и др.). 2. Микроэлементы, содержание которых в клетке составляет 0,000001 - 0,009%. Они представлены преимущественно катионами, участвующими в образовании ферментов, гормонов и других биологически активных веществ (кобальт, медь, цинк, молибден, ванадий, марганец и др.). 3. Ультрамикроэлементы, концентрация которых меньше 0,000001%. К ним отнесены селен, цезий, бериллий, радий, золото и другие редкие элементы, биологическая роль многих из которых еще не выяснена.
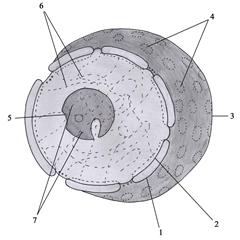
Ядерный аппарат клетки Ядро - главный обязательный органоид клетки у многоклеточных и многих одноклеточных организмов. Впервые ядро в яйцеклетке курицы наблюдал Я. Пуркине в 1825 году. В растительных клетках ядро описано По характеру организации ядерного аппарата живые организмы разделяются на две группы (надцарства): прокариотические и эукариотические. У эукариот ядерный аппарат представлен клеточным ядром, а у прокариот - кольцевой молекулой ДНК. Биологическое значение ядра определяется его главным содержимым - гигантскими молекулами ДНК, способными к транскрипции и трансляции. Основными функциями ядра клетки являются: 1) хранение генетической (наследственной) информации клетки; 2) обеспечение реализации генетической информации, контролирующей разнообразные процессы в клетке; 3) воспроизведение и передача генетической информации при делении клетки. Клетка содержит, как правило, одно ядро, однако существуют и многоядерные клетки. Последние образуются при незавершающемся делении клеток (отсутствует цитотомия или разделение цитоплазмы) либо в результате слияния нескольких одноядерных клеток. Ядро обычно соответствует форме клетки и имеет сферическую форму в сферических или кубических клетках (рис. 8), вытянутую (эллипсовидную) форму в призматических клетках, уплощённую форму в плоских клетках. Однако часто встречаются клетки с бобовидным, палочковидным, многолопастным и сегментированным ядрами. Варьирует и расположение ядра в клетке: оно может располагаться в центре клетки, вблизи её базального (основного) полюса и на периферии под плазмолеммой. Несмотря на постоянство размера ядра для определённого типа клеток, оно может увеличиваться при усилении активности функций клетки и, наоборот, уменьшаться при её ослаблении. Ядро интерфазной (неделящейся) клетки состоит из 4-х компонентов (рис. 21): кариоплазмы (ядерного сока или кариолимфы), хроматина, ядрышка и кариолеммы (ядерной оболочки, или нуклеолеммы). Кариоплазма представляет собой жидкий компонент ядра, состоящий из воды и растворённых (или взвешенных) в ней веществ (РНК, ферменты, гликопротеины, ионы и т.п.). Аналогично гиалоплазме она является бесструктурной фазой, которая создаёт для хроматина и ядрышка специфическое микроокружение, обеспечивающее их нормальное функционирование. Хроматин имеет вид мелких зёрнышек и глыбок, окрашиваемых основными красителями. Он состоит из ДНК и белков. Хроматин являет-

ся, по сути, интерфазным состоянием хромосом и представляет собой длинные и тонкие перекрученные нити, степень спирализации которых различается по их длине. Последнее обстоятельство послужило основанием для выделения двух разновидностей хроматина: эухроматина и гетерохроматина (рис. 21). Эухроматин представляет собой деспирализованные (раскрученные) и участвующие в транскрипции сегменты хромосом. Он плохо окрашивается и практически не виден в световой микроскоп. Гетерохроматин - это спирализованные (плотно скрученные) сегменты хромосом, лишённые активности (не обладающие способностью к транскрипции). Гетерохроматин хорошо окрашивается основными красителями и при наблюдении с помощью светового микроскопа имеет вид гранул и глыбок. Основные скопления гетерохроматина располагаются по периферии ядра, а также вокруг ядрышек. Более мелкие глыбки хроматина разбросаны по всему ядру. Скопление гетерохроматина, соответствующее одной (полностью спирализованной и неактивной) Х-хромосоме у особей женского пола, получило название тельца Барра. Последнее располагается обычно под кариолеммой, а в зернистых лейкоцитах крови имеет вид маленькой добавочной дольки ядра - «барабанной палочки». При повышении синтетической активности клетки, предполагающей интенсификацию процессов транскрипции, количество эухроматина увеличивается, а содержание гетерохроматина уменьшается. Снижение синтетической активности клетки выражается в увеличении содержания гетерохроматина. Полное угнетение функции ядра (например, в повреждённых и гибнущих клетках, в клетках наружных слоёв эпидермиса) ведёт к уменьшению его размеров, исчезновению эухроматина, интенсивному и равномерному окрашиванию основными красителями (явление кариопикноза). Поскольку полностью деспирализованная молекула ДНК (рис. 22) имеет в среднем длину около 5 см, то расположение в ядре диаметром до Первый уровень заключается в образовании нуклеосомной нити: двойная спираль ДНК наматывается на октамер дисковидной формы, как бы дважды обвивая его, и уходит к очередному октамеру. Октамер состоит из 8 глобулярных молекул гистонов (по 2 молекулы каждого из 4-х ти-
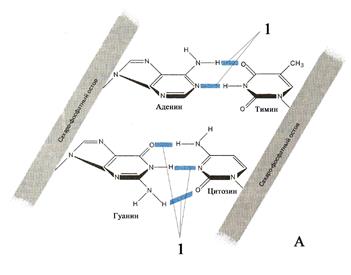
пов белков: Н2А, Н2В, Н3, Н4). Обвитый двумя витками молекулы ДНК октамер назван нуклеосомой (рис. 23). Нуклеосомы разделены короткими участками связывающей (лигандной) ДНК. ДНК хроматина связана также с негистоновыми белками, которые регулируют избирательную активность генов. В такой регуляции могут участвовать и гистоны, ограничивая доступность ДНК для других ДНК-связывающих белков. Второй уровень упаковки приводит к скручиванию нуклеосомной нити в спираль диаметром 30-36 нм, именуемую хроматиновой фибриллой. В период между клеточными делениями каждая хромосома представлена двумя хроматиновыми фибриллами (рис. 24). Третий уровень упаковки ДНК сводится к образованию хроматиновыми фибриллами петель (петельных доменов) диаметром около 300 нм (рис. 24). Четвертый уровень заключается в более плотной упаковке петельных доменов в так называемые «конденсированные участки хромосом» диаметром около 700 нм. Последние являются непосредственной частью метафазной хромосомы (хромосомы, находящейся в метафазе митотического деления) толщиной примерно 1400 нм (рис. 24).
Основной функцией ядрышка является синтез рРНК и сборка её в предшественники субъединиц рибосомы. В процессе транскрипции генов ядрышковых организаторов синтезируется единая крупная молекула рРНК. Последняя связывается с поступившими в ядро из цитоплазмы белками, формируя рибонуклеопротеид (РНП), который затем расщепляется на 3 вида РНК: два из них соединяются с молекулами добавочных белков, образуя предшественники большой субъединицы рибосомы, а третий формирует предшественник малой субъединицы рибосомы. Все предшественники субъединиц поступают через ядерные поры в цитоплазму, где окончательно созревают. Ядрышко обычно окружено перинуклеолярным хроматином, который может частично проникать внутрь его, давая начало интрануклеолярному хроматину. Кариолемма (ядерная оболочка, нуклеолемма) образуется наружной и внутренней мембранами, разделёнными перинуклеарным пространством шириной 15-40 нм (рис. 21). Обе мембраны смыкаются в области ядерных пор. Наружная мембрана кариолеммы является непосредственным продолжением мембран эндоплазматической сети, при этом перинуклеарное пространство соответствует полости цистерн гранулярной эндоплазматической сети (рис. 21). На поверхности наружной мембраны кариолеммы располагаются рибосомы. Внутренняя мембрана кариолеммы гладкая, её интегральные белки связаны с ядерной пластинкой (ламиной) толщиной 80-300 нм. Последняя образована переплетенными промежуточными филаментами, формирующими своеобразный кариоскелет. Ламина выполняет важные функции в поддержании формы ядра, упорядоченной укладке хроматина, формировании кариолеммы при завершении клеточного деления, а также в структурной организации пор. От 3 до 35% поверхности кариолеммы приходится на ядерные поры, количество которых может достигать в клетках животных и человека 2000-4000 (рис. 21). Последние отсутствуют только в ядрах сперматозоидов. С возрастанием активности клетки количество ядерных пор увеличивается. Совокупность структур, связанных с ядерной порой, названа комплексом ядерной поры. В области поры округлые края наружной и внутренней ядерных мембран сомкнуты, а снаружи и изнутри кариолеммы параллельно друг другу расположены два кольца диаметром по 80 нм, каждое из которых содержит 8 белковых гранул (рис. 26). От гранул в центр поры Жизненный цикл клетки Интерфаза Интерфаза занимает не менее 90% времени жизненного цикла клетки. Она включает три периода (рис. 27): постмитотический, или пресинтетический (G1), синтетический (S), премитотический, или постсинтетический (G2). В клеточном цикле существуют так называемые «сверочные точки» (checkpoints), прохождение которых возможно лишь в случае нормального завершения предыдущих этапов и отсутствия поломок. Выделяют по меньшей мере четыре такие точки: точка в периоде G1, точка в периоде S, точка в периоде G2 и «точка проверки сборки веретена деления» в митотическом периоде. Постмитотический период Постмитотический (пресинтетический, G1) период начинается по завершении митотического деления клетки и длится от нескольких часов до нескольких дней. Он характеризуется интенсивным синтезом белка и РНК, увеличением количества органоидов посредством деления или самосборки и, вследствие этого, активным ростом, обусловливающим восстановление нормальных размеров клетки. В течение данного периода синтезируются так называемые «запускающие белки», являющиеся активаторами S-периода. Они обеспечивают достижение клеткой определённого порога (точки рестрикции R), после которого клетка вступает в S-период (рис. 28). Контроль в переходной точке R ограничивает возможность нерегулируемого размножения клеток. Пройдя точку R, клетка переключается на регуляцию внутренними факторами, что обеспечит её митотическое деление.
Премитотический период Премитотический (постсинтетический, G2) период начинается по завершении синтетического периода и продолжается до наступления митоза (рис. 27). Он включает процессы непосредственной подготовки клетки к делению: запасание энергии в АТФ, созревание центриолей, синтез иРНК и белков (в первую очередь тубулина). Продолжительность премитотического периода составляет 2-4 часа (10-20% длительности жизненного цикла). Переход клетки из G2-периода в G0-период, по мнению большинства учёных, невозможен. Вступление клетки в митоз контролируется двумя факторами: Митотический период Митотический период характеризуется протеканием митотического (непрямого) деления клетки, включающего деление ядра (кариокинез) и разделение цитоплазмы (цитокинез). Митоз, занимающий 5-10% времени жизненного цикла и продолжающийся, например, в животной клетке 1-2 часа, подразделяется на четыре основные фазы (рис. 27): профазу, метафазу, анафазу и телофазу. Профаза является самой продолжительной фазой митоза. Она начинается процессом конденсации хромосом (рис. 31), которые обретают, при рассмотрении в световой микроскоп, вид тёмных нитевидных образований. При этом каждая хромосома состоит из двух хроматид, расположенных параллельно и соединенных между собой в области центромеры. Одновременно с конденсацией хромосом происходит диспергация, или распыление ядрышек, которые перестают быть видимыми в световой микроскоп, что связано с вхождением ядрышковых организаторов в состав различных пар хромосом. Соответствующие гены, кодирующие р-РНК, инактивируются. С середины профазы начинает разрушаться кариолемма, распадаясь на фрагменты, а затем на мелкие мембранные пузырьки. Гранулярная эндоплазматическая сеть распадается на короткие цистерны и вакуоли, на мембранах которых резко уменьшается количество рибосом. Примерно на четверть уменьшается число полисом, локализованных как на мембранах, так и в гиалоплазме клетки. Такие изменения приводят к резкому падению уровня синтеза белка в делящейся клетке. Важнейшим процессом профазы является формирование митотического веретена. Репродуцировавшиеся ещё в S-периоде центриоли начинают расходиться к противоположным концам клетки, где впоследствии сформируются полюсы веретена. К каждому полюсу перемещается диплосома (две центриоли). Одновременно формируются микротрубочки, отходящие от одной центриоли каждой диплосомы (рис. 32). Формирующееся в результате этого образование имеет в животной клетке веретеновидную форму, в связи с чем получило название «веретена деления» клетки. Оно состоит из трёх зон: двух зон центросфер с центриолями внутри них и

располагающейся между ними зоны нитей веретена деления. Все три зоны содержат большое количество микротрубочек. Последние входят в состав центросфер, располагаясь вокруг центриолей, формируют нити ве 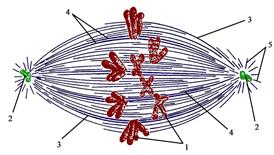 ретена, а также подходят к центромерам хромосом (рис. 33). Микротрубочки, тянущиеся от одного полюса к другому (не прикрепляющиеся к центромерам хромосом), получили название полюсных микротрубочек. Микротрубочки, отходящие от кинетохо ретена, а также подходят к центромерам хромосом (рис. 33). Микротрубочки, тянущиеся от одного полюса к другому (не прикрепляющиеся к центромерам хромосом), получили название полюсных микротрубочек. Микротрубочки, отходящие от кинетохо  ров (центромер) каждой хромосомы к полюсу веретена, названы кинетохорными микротрубочками (нитями). Входящие в состав центросфер и лежащие вне веретена деления микротрубочки, ориентированные от центриолей к плазмолемме названы астральными микротрубочками, или микротрубочками сияния (рис. 33). Все микротрубочки веретена находятся в динамическом равновесии между сборкой и разборкой. При этом около 108 молекул тубулина организованы в микротрубочки. Центромеры (кинетохоры) сами способны индуцировать сборку микротрубочек. Следовательно, центриоли и хромосомные центромеры являются в животной клетке центрами организации микротрубочек веретена деления. В индукции роста микротрубочек в зоне полюса деления принимает участие только одна (материнская) центриоль. ров (центромер) каждой хромосомы к полюсу веретена, названы кинетохорными микротрубочками (нитями). Входящие в состав центросфер и лежащие вне веретена деления микротрубочки, ориентированные от центриолей к плазмолемме названы астральными микротрубочками, или микротрубочками сияния (рис. 33). Все микротрубочки веретена находятся в динамическом равновесии между сборкой и разборкой. При этом около 108 молекул тубулина организованы в микротрубочки. Центромеры (кинетохоры) сами способны индуцировать сборку микротрубочек. Следовательно, центриоли и хромосомные центромеры являются в животной клетке центрами организации микротрубочек веретена деления. В индукции роста микротрубочек в зоне полюса деления принимает участие только одна (материнская) центриоль.
Метафаза занимает около трети времени всего митоза. В течение этой фазы заканчивается образование веретена деления и достигается максимальный уровень конденсации хромосом. Последние выстраиваются в области экватора митотического веретена (рис. 31, 34), формируя так называемую «метафазную (экваториальную) пластинку» (вид сбоку) или «материнскую звезду» (вид со стороны полюса клетки). Хромосомы удерживаются в экваториальной плоскости благодаря сбалансированному натяжению центромерных (кинетохорных) микротрубочек. К концу метафазы завершается обособление сестринских хроматид: их плечи лежат параллельно друг другу, а между ними видна разделяющая их щель. Последним местом контакта между хроматидами остаётся центромера. Анафаза является самой короткой фазой, занимающей лишь несколько процентов времени митоза. Она начинается утратой связи между сестринскими хроматидами в области центромер и движением хро-
В телофазе - конечной стадии митоза - вокруг каждой полюсной группы хромосом (дочерние звёзды) образуется ядерная оболочка: фрагменты кариолеммы (мембранные пузырьки) связываются с поверхностью отдельных хромосом, частично окружают каждую из них и только после этого сливаются, образуя полную ядерную оболочку (рис. 31, 34). После восстановления ядерной оболочки возобновляется синтез РНК, из соответствующих участков (ядрышковых организаторов) хромосом оформляется ядрышко и деконденсируется хроматин, переходя в типичное для интерфазы дисперсное состояние. Ядра клеток постепенно увеличиваются, а хромосомы прогрессивно деспирализуются и исчезают. Одновременно углубляется клеточная перетяжка, а соединяющий их цитоплазматический мостик с пучком микротрубочек внутри сужается (рис. 31). Последующая перешнуровка цитоплазмы завершает разделение цитоплазмы (цитокинез). Равномерному разделению органелл между дочерними клетками способствует их большое количество в клетке (митохондрии) либо распад во время митоза на мелкие фрагменты и мембранные пузырьки. При повреждении веретена деления может происходить атипический митоз, ведущий к неравномерному распределению генетического материала между клетками (анэуплоидия). Отдельные атипические митозы, при которых цитотомия отсутствует, завершаются образованием гигантских клеток. Атипичные митозы свойственны обычно клеткам злокачественных опухолей и облучённых тканей. Существует вариант митотического деления, при котором ядерная оболочка не разрушается и веретено деления не формируется. Удвоившийся набор хромосом остаётся в одном ядре. Такое деление клетки названо эндомитозом. Повторные эндомитозы ведут к полиплоидии - значительному увеличению числа хромосом в ядре. Полиплоидия может быть результатом обычных незавершённых митозов. Полиплоидные клетки отличаются повышенной функциональной активностью. Дистрофия клетки
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-02-19; просмотров: 144; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.218.172.249 (0.071 с.) |


 Впервые растительные клетки, а точнее, оболочки мёртвых клеток в срезах пробки (рис. 1) описаны английским естествоиспытателем Робертом Гуком. Результаты описания были представлены им на заседании Королевского общества естествоиспытателей в Лондоне в 1655 году. Р. Гук показал, что всё вещество пробки состоит из большого числа маленьких отделений или полостей, наполненных воздухом и разграниченных тонкими перегородками. Эти полости, или ячейки, он назвал «клетками». Термин «клетка» утвердился и сохранился в биологии до настоящего времени, несмотря на то, что Роберт Гук наблюдал, собственно, не клетки, а лишь сохранившиеся от них целлюлозные оболочки и что клетки оказались далеко не полостями.
Впервые растительные клетки, а точнее, оболочки мёртвых клеток в срезах пробки (рис. 1) описаны английским естествоиспытателем Робертом Гуком. Результаты описания были представлены им на заседании Королевского общества естествоиспытателей в Лондоне в 1655 году. Р. Гук показал, что всё вещество пробки состоит из большого числа маленьких отделений или полостей, наполненных воздухом и разграниченных тонкими перегородками. Эти полости, или ячейки, он назвал «клетками». Термин «клетка» утвердился и сохранился в биологии до настоящего времени, несмотря на то, что Роберт Гук наблюдал, собственно, не клетки, а лишь сохранившиеся от них целлюлозные оболочки и что клетки оказались далеко не полостями. В дальнейшем клеточное строение многих частей растений подтвердили и описали М. Мальпиги (1675) и Н. Грю (1671). Клетки животных (эритроциты, сперматозоиды), а также одноклеточные организмы впервые увидел А. Левенгук (1674).
В дальнейшем клеточное строение многих частей растений подтвердили и описали М. Мальпиги (1675) и Н. Грю (1671). Клетки животных (эритроциты, сперматозоиды), а также одноклеточные организмы впервые увидел А. Левенгук (1674). Однако от первых описаний клеток до формулирования клеточной теории прошло более полутора веков - период такой же длительный, как и вся современная история развития цитологии (с 1839 года по настоящее время). Столь медленные темпы развития науки о клетке в период её предыстории объяснялись несовершенством первых микроскопов, отличавшихся высокой хроматической аберрацией и обусловленной ею нечёткостью изображения, а также отсутствием специальных методов подготовки биологических объектов к микроскопическим исследованиям.
Однако от первых описаний клеток до формулирования клеточной теории прошло более полутора веков - период такой же длительный, как и вся современная история развития цитологии (с 1839 года по настоящее время). Столь медленные темпы развития науки о клетке в период её предыстории объяснялись несовершенством первых микроскопов, отличавшихся высокой хроматической аберрацией и обусловленной ею нечёткостью изображения, а также отсутствием специальных методов подготовки биологических объектов к микроскопическим исследованиям. всех организмов. Особое значение имели исследования, проведённые чешским учёным Я. Пуркинье и сотрудниками берлинской лаборатории И. Мюллера. В 1838 году вышла в свет работа немецкого ботаника М. Шлейдена «Материалы к фитогенезу», в которой он показал, что клетка является основной структурной единицей растений и поставил вопрос о способе образования новых клеток. Последний, однако, М. Шлейден решил довольно примитивно,
всех организмов. Особое значение имели исследования, проведённые чешским учёным Я. Пуркинье и сотрудниками берлинской лаборатории И. Мюллера. В 1838 году вышла в свет работа немецкого ботаника М. Шлейдена «Материалы к фитогенезу», в которой он показал, что клетка является основной структурной единицей растений и поставил вопрос о способе образования новых клеток. Последний, однако, М. Шлейден решил довольно примитивно,  немецкий зоолог Т. Шванн в книге «Микроскопические
немецкий зоолог Т. Шванн в книге «Микроскопические  Основные положения клеточной теории Шлейдена-Шванна заключаются в следующем:
Основные положения клеточной теории Шлейдена-Шванна заключаются в следующем:



 Ядрышко выявляется при световой микроскопии как мелкая плотная гранула диаметром 1-3 мкм. Ядрышко отличается высокой концентрацией рибонуклеопротеида, в связи с чем интенсивно окрашивается основными красителями. С повышением функциональной активности клетки увеличиваются как размеры, так и количество ядрышек. Наиболее крупные ядрышки характерны для эмбриональных и активно синтезирующих белки клеток, а также для клеток быстрорастущих злокачественных опухолей.
Ядрышко выявляется при световой микроскопии как мелкая плотная гранула диаметром 1-3 мкм. Ядрышко отличается высокой концентрацией рибонуклеопротеида, в связи с чем интенсивно окрашивается основными красителями. С повышением функциональной активности клетки увеличиваются как размеры, так и количество ядрышек. Наиболее крупные ядрышки характерны для эмбриональных и активно синтезирующих белки клеток, а также для клеток быстрорастущих злокачественных опухолей. Ядрышко формируется участками различных молекул ДНК, кодирующих рибосомную РНК (рРНК). Такие участки ДНК названы «ядрышковыми организаторами» (рис. 25). У человека они находятся в ДНК 13й, 14й, 15й, 21й и 22й пар хромосом. Ядрышко не имеет собственной оболочки (мембраны) и исчезает (диспергируется) в профазе митоза из-за того, что ядрышковые организаторы «растаскиваются» в соответствующие пары формирующихся хромосом. В телофазе митотического деления ядрышки формируются вновь участками деконденсирующихся хромосом.
Ядрышко формируется участками различных молекул ДНК, кодирующих рибосомную РНК (рРНК). Такие участки ДНК названы «ядрышковыми организаторами» (рис. 25). У человека они находятся в ДНК 13й, 14й, 15й, 21й и 22й пар хромосом. Ядрышко не имеет собственной оболочки (мембраны) и исчезает (диспергируется) в профазе митоза из-за того, что ядрышковые организаторы «растаскиваются» в соответствующие пары формирующихся хромосом. В телофазе митотического деления ядрышки формируются вновь участками деконденсирующихся хромосом. простираются фибриллы толщиной около 5 нм, сходящиеся у центральной гранулы (по мнению некоторых учёных, она представляет собой транс-
простираются фибриллы толщиной около 5 нм, сходящиеся у центральной гранулы (по мнению некоторых учёных, она представляет собой транс- центральной гранулы. Ядерные по-
центральной гранулы. Ядерные по-
 Клетка может не достигнуть точки R и выйти из клеточного цикла, вступив в период репродуктивного покоя (G0). Причинами такого выхода могут быть: 1) необходимость дифференцироваться и выполнять специфические функции; 2) потребность преодолеть период неблагоприятных условий или вредных воздействий среды; 3) необходимость восстановить повреждённую ДНК. Из периода репродуктивного покоя (G0) одни клетки могут возвращаться в клеточный цикл, а другие утрачивают эту способность в ходе дифференцировки. В связи с этим понадобился безопасный момент прекращения прохождения клеточного цикла, которым и стала точка R. Предполагается, что механизм регуляции клеточного роста, включающий специфическую точку R, мог возникнуть из-за условий существования или взаимодействия с другими клетками, требующими прекращения деления. Про клетки, остановленные в этом покоящемся состоянии, говорят, что они вступили в фазу G0 клеточного цикла.
Клетка может не достигнуть точки R и выйти из клеточного цикла, вступив в период репродуктивного покоя (G0). Причинами такого выхода могут быть: 1) необходимость дифференцироваться и выполнять специфические функции; 2) потребность преодолеть период неблагоприятных условий или вредных воздействий среды; 3) необходимость восстановить повреждённую ДНК. Из периода репродуктивного покоя (G0) одни клетки могут возвращаться в клеточный цикл, а другие утрачивают эту способность в ходе дифференцировки. В связи с этим понадобился безопасный момент прекращения прохождения клеточного цикла, которым и стала точка R. Предполагается, что механизм регуляции клеточного роста, включающий специфическую точку R, мог возникнуть из-за условий существования или взаимодействия с другими клетками, требующими прекращения деления. Про клетки, остановленные в этом покоящемся состоянии, говорят, что они вступили в фазу G0 клеточного цикла.
 Движение хромосом складывается из двух процессов: расхождения их по направлению к полюсам и дополнительного расхождения самих полюсов. Предположения о сокращении (саморазборке) микротрубочек как о механизме расхождения хромосом в митозе не подтвердились. Поэтому многие исследователи поддерживают гипотезу «скользящих нитей», согласно которой соседние микротрубочки, взаимодействуя друг с другом (например, хромосомные и полюсные) и с сократительными белками (миозин, динеин), тянут хромосомы к полюсам.
Движение хромосом складывается из двух процессов: расхождения их по направлению к полюсам и дополнительного расхождения самих полюсов. Предположения о сокращении (саморазборке) микротрубочек как о механизме расхождения хромосом в митозе не подтвердились. Поэтому многие исследователи поддерживают гипотезу «скользящих нитей», согласно которой соседние микротрубочки, взаимодействуя друг с другом (например, хромосомные и полюсные) и с сократительными белками (миозин, динеин), тянут хромосомы к полюсам. Анафаза завершается скоплением на полюсах клетки по одному, идентичному друг другу, набору хромосом, формирующему так называемую «дочернюю звезду». В конце анафазы в животной клетке начинает образовываться клеточная перетяжка, углубляющаяся в следующей фазе и приводящая к цитотомии (цитокинезу). В её образовании участвуют актиновые миофиламенты, концентрирующиеся по окружности клетки в виде «сократимого кольца».
Анафаза завершается скоплением на полюсах клетки по одному, идентичному друг другу, набору хромосом, формирующему так называемую «дочернюю звезду». В конце анафазы в животной клетке начинает образовываться клеточная перетяжка, углубляющаяся в следующей фазе и приводящая к цитотомии (цитокинезу). В её образовании участвуют актиновые миофиламенты, концентрирующиеся по окружности клетки в виде «сократимого кольца».


