Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Водорості: характеристика, особливості біології, значення для людиниСодержание книги
Поиск на нашем сайте
Значення водоростей у природі та для людини. Значення водоростей у народному господарстві. Водорості широко використовують у народному господарстві. Багато з них людина здавна використовувала в їжу (ламінарію, порфіру), на корм худобі, як добрива. Діатоміти використовують у харчовій, хімічній, фармацевтичній промисловості, будівництві. Бурі водорості є сировиною для добування альгінатів (солей альгінової кислоти), які застосовують у різних галузях народного господарства. Так, на основі альгінату натрію виготовляють клей, який використовують у текстильному виробництві, для проклеювання паперу, скріплення цементу. Плівки з альгінату натрію, нанесені на бетонні споруди, метали, верстати, деревину, захищають їх від корозії, гниття, руйнування. З червоних водоростей (філофора) добувають агар, який використовують у мікробіології та кондитерській промисловості, із зелених (кладофора, ризоклоніум) виготовляють папір. Деякі види водоростей мають цілющі властивості і використовуються в медицині (ламінарії, лікувальні грязі з синьозеленими водоростями; добування йоду). Багато водоростей є біоіндикаторами під час санітарно-біологічної оцінки вод або виконують функцію активних санітарів забруднених водойм. У деяких країнах водорості вирощують у штучних водоймах для промислового виробництва органічних речовин. Разом з тим водорості можуть мати і негативне значення. Так, у разі масового розмноження у водоймах вони спричинюють "цвітіння" води, роблячи її непридатною для використання, забруднюють насосні станції та водоводи, деякі види вкривають днища суден, буї, погіршуючи їх експлуатацію. Амебоїдний (ризоподіальний) Одноклітинні організми, позбавлені твердої клітинної оболонки й внаслідок цього, не здатні зберігати постійну форму тіла. Завдяки відсутності клітинної стінки й наявності особливих внутрішньоклітинних структур клітина здатна до плазуючого руху за допомогою псевдоподій або ризоподій. Для деяких видів характерний утвір багатоядерного плазмодія шляхом злиття декількох амебоїдних клітин. Амебоїдне будову можуть удруге здобувати деякі монадні форми шляхом відкидання або втягування джгутиків. 2 Монадний (мал. 3) Одноклітинні водорості, що мають постійну форму тіла, джгутики, часто стигму, а прісноводні - скорочувальну вакуоль. Клітини активно рухаються у вегетативному стані. Часто зустрічається об'єднання декількох монадних клітин у колонію, оточену загальним слизом, у деяких випадках навіть з'єднаних між собою за допомогою плазмодесм. У високоорганізованих форм із багатоклітинним талломом часто є стадії розсілення - зооспори й гамети, що мають монадную структуру.
Коккоїдний Одноклітинні, позбавлені яких-небудь органоїдів пересування, що й зберігають постійну форму тіла у вегетативному стані клітини. Найчастіше є стовщена клітинна стінка або панцир, можуть бути різні вирости, пори та ін. для полегшення ширяння в товщі води. Багатьом водоростям з даною структурою властивий утвір колоній. Деякі діатомеї й десмідієві здатні до активного пересування шляхом виділення слизу. Пальмеллоїдний (капсульний) Постійний, досить великий, як правило, прикріплений до субстрату, утвір з декількох коккоїдних клітин, занурених у загальну слизувату масу. Клітини безпосередньо між собою не поєднуються - відсутні плазмодесми. Тимчасову стадію життєвого циклу з аналогічною морфологією називають пальмеллевидним станом. У такий стан можуть переходити багато монадних й коккоїдних водоростей при настанні несприятливих умов.
5 Нитчатий (мал. 4)
Клітки з'єднані в нитку, просту або розгалужену. Нитки можуть вільно плавати в товщі води, прикріплюватися до субстрату, або поєднуватися в колонію. Вегетативно нитчаті водорості розмножуються звичайно розпадом нитки на окремі фрагменти. Ріст ниток може йти чотирма шляхами: дифузійним - діляться всі клітини нитки, інтеркалярним - зона росту розташована в середині нитки, апікальним - розподілом кінцевих клітин, і базальним - розподілом клітин уоснові таллома. Клітини в нитці не мають джгутиків і можуть бути зв'язані між собою плазмодесмами. Пластинчастий Багатоклітинні талломы у формі пластинок з одного, двох або декількох шарів клітин. Виникають при поздовжньому розподілі клітин, щоскладають нитку. Число шарів залежить від характеру утвору перегородок при розподілі клітин. Іноді шари можуть розходитися, і таллом тоді здобуває трубчасту форму (порожній у середині), стінки при цьому стають одношаровими.
Сифонокладальний Таллом представлений багатодерними клітинами, з'єднаними в нитчаті або іншої форми багатоклітинні талломы(Siphonocladales). Сарциноїдний Колонії, що представляють собою групи (пачки або ниткоподібні утворення), які виникають у результаті розподілу однієї вихідної клітини й включеннях, що розтягують оболонку цієї клітини.
Псевдопаренхіматозний Представлений сланями, які утворювалися в результаті зрощення розгалужених ниток, нерідко супроводжуваний морфофункціональною диференціацією, що має вигляд неправильних тканин.
Список літератури 1. Урок на тему: "Многообразие водорослей" автор Яковлева Л. А. 2. М.М. Мусієнко, П.С. Славний, П.Г. Балан, Біологія, 7 клас 3. Тимонин А.К. Ботаника. Том 4. Систематика высших растений. Книга 2 Учебник для студ. высш. учеб. заведений. Издательский центр "Академия". 2009. -- 352 с. 4. Ботаника. Андреева И.И., Родман Л.С. 2 - е изд., перераб. и доп. - М.: КолосС, 2002. - 488 с. 5. Биология. Современная иллюстрированная энциклопедия л. ред. Горкин А. П. - М.: Росмэн-Пресс, 2006. - 560 с. (Серия: Современная иллюстрированная энциклопедия.)
Водорості: характеристика, особливості біології, значення для людини План 1. Загальна характеристика нижчих рослин – водоростей. 2.Різноманітність бурих водоростей 3. Значення водоростей у природі та житті людини. Розмноження водоростей 5.Різноманітність бурих водоростей 1. Загальна характеристика нижчих рослин – водоростей. Загальна характеристика. Водорості — велика група найдавніших рослин. Будова їхнього тіла і розміри характеризуються значною різноманітністю. Існують одноклітинні, багатоклітинні і колоніальні форми мікроскопічних розмірів (від тисячних часток міліметра), а також форми з різною будовою слані, що досягають 30—45 м. Водорості — єдина група організмів, серед яких зустрічаються прокаріоти (синьозелені) і еукаріоти (решта відділів). В ядрах еукаріотних водоростей виявлені структури, властиві ядрам інших еукаріотів: оболонки, ядерний сік, ядерця, хромосоми. Загальною ознакою всіх водоростей є наявність хлорофілу. Крім хлорофілу водорості можуть містити й інші пігменти (фікоціан, фікоеритрин, каротин, ксантофіл, фіко-сантин). Щ пігменти надають водоростям червоного, бурого, жовто-зеленого кольору, маскуючи основний зелений. Наявність пігментів у клітинах водоростей забезпечує автотрофний тип живлення. Проте багато водоростей здатні за певних умов переходити на гетеротрофне живлення (евгленові — в темряві) або поєднувати його з фотосинтезом (міксотрофний тип живлення). Кількість видів водоростей перевищує 40 тис. Проте класифікація їх не завершена, оскільки не всі форми достатньо вивчені. У нашій країні прийнято поділяти водорості на 10 відділів: синьозелені, пірофітові, золотисті, діатомові, жовтозелені, бурі, червоні, евгленові, зелені, харові. Найбільшу кількість видів налічують зелені (13—20 тис.) і діатомові (10 тис.) водорості. Поділ водоростей на відділи збігається зазвичай з їхнім забарвленням, яке, як правило, пов'язане з особливостями будови клітин і слані. Будова, склад і властивості клітинних компонентів водоростей характеризуються великою різноманітністю. У процесі еволюції природний добір зберіг найперспек-тивніші форми, в тому числі такий тип клітинної організації, який дав змогу рослинам перейти до наземного способу життя. Розмноження водоростей буває вегетативним, безстатевим (за допомогою спор) і статевим. У одного й того самого виду залежно від умов і пори року способи розмноження різні. При цьому спостерігається зміна ядерних фаз — гаплоїдної і диплоїдної. Сприятливими умовами для зростання водоростей є наявність світла, джерел вуглецю та мінеральних солей, а основним середовищем життя для них є вода.Значний вплив на життя водоростей мають температура, солоність води тощо. За місцем зростання водорості поділяють на дві великі групи: ті, що живуть у водоймах, та ті, що живуть поза водоймами. Серед тих, що живуть у водоймах, виділяють такі екологічні групи: планктон — дуже дрібні водорості, які знаходяться в товщі води у завислому стані (хламідомонада, вольвокс, пандорина, мікроцистіс); бентос — водорості, що живуть на дні водойм (харові, багато діатомових); перифітон — водорості, якими обростають підводні предмети або вищі рослини водойм (кладофора, червоні та бурі водорості морів). Населяють водорості і гарячі джерела, а також водойми з підвищеною концентрацією солей. Водорості, що живуть поза водоймами, також поділяють на групи: грунтові (едафітон) — живуть у грунті або на грунті (ботридій, деякі вошерії), у грунті живе понад 700 видів водоростей з різних відділів; наземні (аерофі-тон) — на корі дерев, на скелях (трентеполія, плеврокок). Багато водоростей вступають у симбіотичні зв'язки з іншими представниками рослинного й тваринного світу. Особливе місце тут належить симбіозу водоростей з грибами. У цьому симбіозі виникає така біологічна єдність двох організмів, що призводить до появи третього — ли-шайника, який відрізняється і від першого, і від другого. Зелені водорості. Ознайомимося з представниками відділу зелених водоростей, розглянувши з одноклітинних хламідомонаду, плеврокок і хлорелу, з багатоклітинних нитчастих — улотрикс і спірогіру. Хламідомонада — мікроскопічна одноклітинна водорість грушоподібної або овальної форми. Поверхня клітини вкрита прозорою безбарвною пектиновою оболонкою. На передньому кінці тіла оболонка утворює невелике випинання — носик, від якого відходять два джгутики. За допомогою цих джгутиків водорість рухається. Всю внутрішню частину клітини займає цитоплазма з великим чашоподібним хлоропластом (хроматофором) зеленого кольору. В нижній потовщеній частині хлоропласта розміщений кулястий піреноїд (зона, де найактивніше синтезуються й нагромаджуються поживні речовини), який містить багато білків і оточений зернами крохмалю. На верхній частині хроматофора знаходиться добре помітне червоне вічко, насичене каротином. Функція його невідома. В заглибині хроматофора розміщене велике кулясте ядро з добре помітним ядерцем. Спереду біля основи джгутиків розміщені дві пульсівні вакуолі. Хламідомонади розмножуються статевим і безстатевим шляхом (мал. 62). При безстатевому розмноженні клітина втрачає джгутики, її ядро, хлоропласт і цитоплазма Діляться на 4 (рідше 8) клітин — зооспор. У кожної дочірньої клітини виростає по 2 джгутики, оболонка материнської клітини руйнується і зооспори виходять у воду. У такий спосіб водорості розмножуються дуже швидко. Же через добу дочірні клітини знов діляться. При статевому розмноженні в материнській клітині утворюються гамети. Вони подібні до зооспор, однак кількість значно більша — 32 або 64 в одній клітині. Після дозрівання гамети виходять з материнської клітини і попарно з'єднуються, утворюючи зиготу. Вона вкривається исною оболонкою і переходить у стан спокою. Через який час зигота виходить з оболонки і ділиться мейочно з утворенням 4 гаплоїдних зооспор.
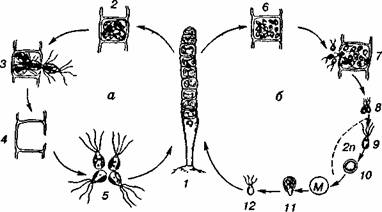
2. Мал.1 Загальна характеристика нижчих рослин – водоростей. Особливості біології зелених водоростей. . Життєвий цикл хламідомонади: а — безстатеве розмноження; б — статеве розмноження; М ~ мейоз; 1 — вегетативна особина; 2 — утворення зооспор; 3 — зооспори; 4 — утворення гамет; 5 — копуляція гамет; 6 — зигота; 7 — проростання зиготи; 8 — зооспора Хламідомонади живуть у невеликих, дуже забруднених водоймах, що добре прогріваються (калюжі, стічні канави). У таких водоймах вони дуже швидко розмножуються і спричинюють "цвітіння" води. Оскільки поряд з автотрофним способом живлення клітини хламідомонад поглинають розчинені органічні речовини, вони тим самим сприяють процесу очищення забрудненої води (самоочищенню). Цей процес можна спостерігати у відстійниках, очисних спорудах міської каналізації. Уже через кілька днів після чергового викидання стічних вод водойма очищається. Вода стає чистою і прозорою до дна. Водоростей — активних санітарів — уже немає в товщі води, їхні зиготи опустились на дно і там "відпочивають" до чергового забруднення, яке знов їх активізує. Плеврокок — мікроскопічна одноклітинна наземна водорість без джгутиків. Під щільною безбарвною оболонкою клітини знаходяться цитоплазма, ядро і пластинчастий хлоропласт. Зазвичай клітини поєднані в групи по 4—6 і більше у вигляді пакетів, іноді утворюють короткі нитки. Пакети можуть розпадатися на окремі клітини» кожна з них існує самостійно і швидко починає ділитися. Клітини плеврокока діляться в двох взаємно перпендикулярних напрямках, утворюючи нові пакети. Статевого розмноження немає, зооспори не утворюються. Поселяється плеврокок на корі старих дерев, особливо на північному боці на землі в сирих місцях, на стінках глиняних горщиків з кімнатними рослинами, утворюючи на них разом з іншими водоростями зелений наліт. Плеврокок входить до складу слані багатьох лишайників. Хлорела — одноклітинна зелена водорість, має вигляд мікроскопічної нерухомої (без джгутиків) кульки до. 15 мкм у діаметрі. Зовні клітини вкриті твердою двоконтурною оболонкою целюлозної природи. В цитоплазмі міститься один чашоподібний хлоропласт з одним піре-ноїдом у потовщеній його частині. Ядро одне, однак у живій клітині без спеціальної обробки його не видно. Розмножується лише нестатевим шляхом. При цьому в материнській клітині утворюється 4—8 апланоспор, які через розрив оболонки виходять у воду й набувають вигляду дорослої особини. Хлорела невибаглива до умов існування і здатна до інтенсивного розмноження, тому зустрічається всюди: у прісних водоймах, морях і грунті. Вона може вступати у симбіоз з різними організмами, входить до складу лишайників. Клітина хлорели — зручний об'єкт для різних досліджень. Хлорела — основний об'єкт масового культивування водоростей для практичного використання в різних напрямах. Значну роль у формуванні підвищеного інтересу до неї відіграв її хімічний склад. У перерахунку на суху речовину хлорела містить повноцінних білків 40 % і більше, ліпідів — до 20 %, вуглеводів — до 35 %, зольних речовин — до 10 %. Є вітаміни групи В, аскорбінова кислота (віт. С) і філохінони (віт. К). Знайдено речовину, яка має антибіотичну активність — "хлорелін". У деяких країнах хлорелу використовують у їжу після спеціальної обробки, Що поліпшує її засвоєння. Для споживання використовують свіжу біомасу хлорели або спеціальну пасту з неї. Крім одноклітинних форм до відділу зелених водоростей належать і багатоклітинні нитчасті. Форма їхнього тіла має вигляд простих або розгалужених ниток, що безперервно ростуть у довжину в результаті поділу клітин упоперек. Найтиповішими представниками нитчастих водоростей є улотрикс і спірогіра. Улотрикс дуже поширений у річках, живе, прикріплюючись до підводних предметів, утворюючи яскраво-зелені ростання. Баговиння улотрикса складається з нерозгалужених ниток різної довжини, які на початку росту прикріплюються до субстрату безбарвною видовженою клітиною — ризоїдом. Клітини ниток циліндричні або бочкоподібні, короткі. Кожна клітина має ядро, пристінний хлоропласт у вигляді неповного кільця і один або кілька піреноїдів. Розмноження вегетативне, безстатеве і статеве (мал. 2). При вегетативному розмноженні нитка улотрикса розпадається на короткі сегменти, кожний з яких росте, утворюючи нову нитку. Безстатеве розмноження здійснюється за допомогою зооспор, які формуються в клітинах. Зооспори — яйце-або грушоподібні клітини з чотирма джгутиками на передньому кінці, червоним вічком і двома пульсівними вакуолями. Кожна зооспора, вийшовши назовні, через деякий час прикріплюється до субстрату і проростає в дорослу особину. При статевому розмноженні в клітинах нитки формуються гамети, подібні до зооспор, однак лише з двома джгутиками. Гамет у кожній клітині більше, ніж зооспор. З'єднуючись попарно, гамети однієї й тієї самої або частіше різних ниток утворюють зиготу. Вона залишається рухливою недовго, скоро осідає на дно водойми, втрачає джгутики і вкривається товстою оболонкою, що захищає ЇЇ від несприятливих умов. Під час проростання зиготи відбувається редукційний поділ з утворенням 4—16 гаплоїдних особин.
Мал. 2. Життєвий цикл улотрикса: а — безстатеве розмноження; б — статеве розмноження; М — мейоз; / — вегетативна особина; 2 — утворення зооспор; 3 — вихід зооспор; 4 — порожня клітина; 5 — зооспори; 6 — утворення гамет; 7 — вихід гамет; 8 — ізогамія; 9, 10 — зиготи; 11 — проростання зиготи; 12 — зооспора Спірогіра — одна з найпоширеніших зелених нитчастих водоростей у прісноводних басейнах. Довгі нитки слані утворюють сплетення (баговиння) яскраво-зеленого кольору, до субстрату вони не прикріплюються і вільно плавають у воді. Нитки спірогіри завдовжки від кількох міліметрів до 8-10 см не галузяться і складаються з одного ряду однакових видовжених циліндричних клітин. Кожна клітина має двошарову оболонку, яка зовні вкрита слизовим чохлом. Внутрішній шар оболонки складається з целюлози, зовнішній — з пектинових речовин. Целюлозна оболонка оточує цитоплазму, в якій розміщені спіральне закручені зелені стрічкоподібні хлоропласти з численними піреноїдами. Велике ядро з добре помітним ядерцем розміщене в центральній частині клітини, оточене шаром цитоплазми, який сполучений з пристінним шаром тяжами. Проміжки між тяжами заповнені вакуолями. Спірогіра розмножується вегетативне і статевим шляхом, спор вона ніколи не утворює. Вегетативне розмноження відбувається в разі випадкового розриву ниток або розпадання її на окремі клітини за несприятливих умов. З кожної частини нитки або окремої клітини утворюються нові особини поділом клітин. Статеве розмноження здійснюється кон'югацією. При цьому дві нитки, зовні подібні, зближуються. З клітин ниток, розміщених одна навпроти одної, утворюються вирости. При стиканні цих виростів (мал. 3) перегородки їх зникають і вміст "чоловічої" клітини переливається в "жіночу". В результаті цього злиття утворюється зигота, яка вкривається товстою оболонкою і після певного періоду спокою проростає. При цьому вона ділиться мейозом, утворюючи чотири гаплоїдні клітини. Три з них — дрібні — відмирають, а одна — велика — разом із вмістом зиготи утворює проросток нової особини, який дає початок новій нитці. Розмноження
|
||||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 208; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.148.117.240 (0.014 с.) |